Mycobacterium tuberculosis Rv2005c Induces Dendritic Cell Maturation and Th1 Responses and Exhibits Immunotherapeutic Activity by Fusion with the Rv2882c Protein


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
2.1. Mice
2.2. Infection, Bacterial Strains and Cell Preparations
2.3. Purification of Recombinant Protein
2.4. Cytotoxicity Analysis
2.5. Enzyme-Linked Immunosorbent Assay (ELISA)
2.6. Flow Cytometry Analysis
2.7. Western Blot Analysis
2.8. In Vitro T Cell Proliferation Assay
2.9. Hypoxia Induced in Bmdms by Cocl2
2.9.1. Cytotoxicity Analysis
2.9.2. Western Blot Analysis
2.9.3. Hypoxia-induced Intracellular Survival of Mtb
2.10. Vaccine Evaluation
2.10.1. In Vivo Chemotherapy Assay
2.10.2. Analysis of Antigen-Specific T Cell Cytokine Secretion
2.10.3. Intracellular Cytokine Assays
2.10.4. Bacterial Count Analysis
2.11. Statistical Analysis
3. Results
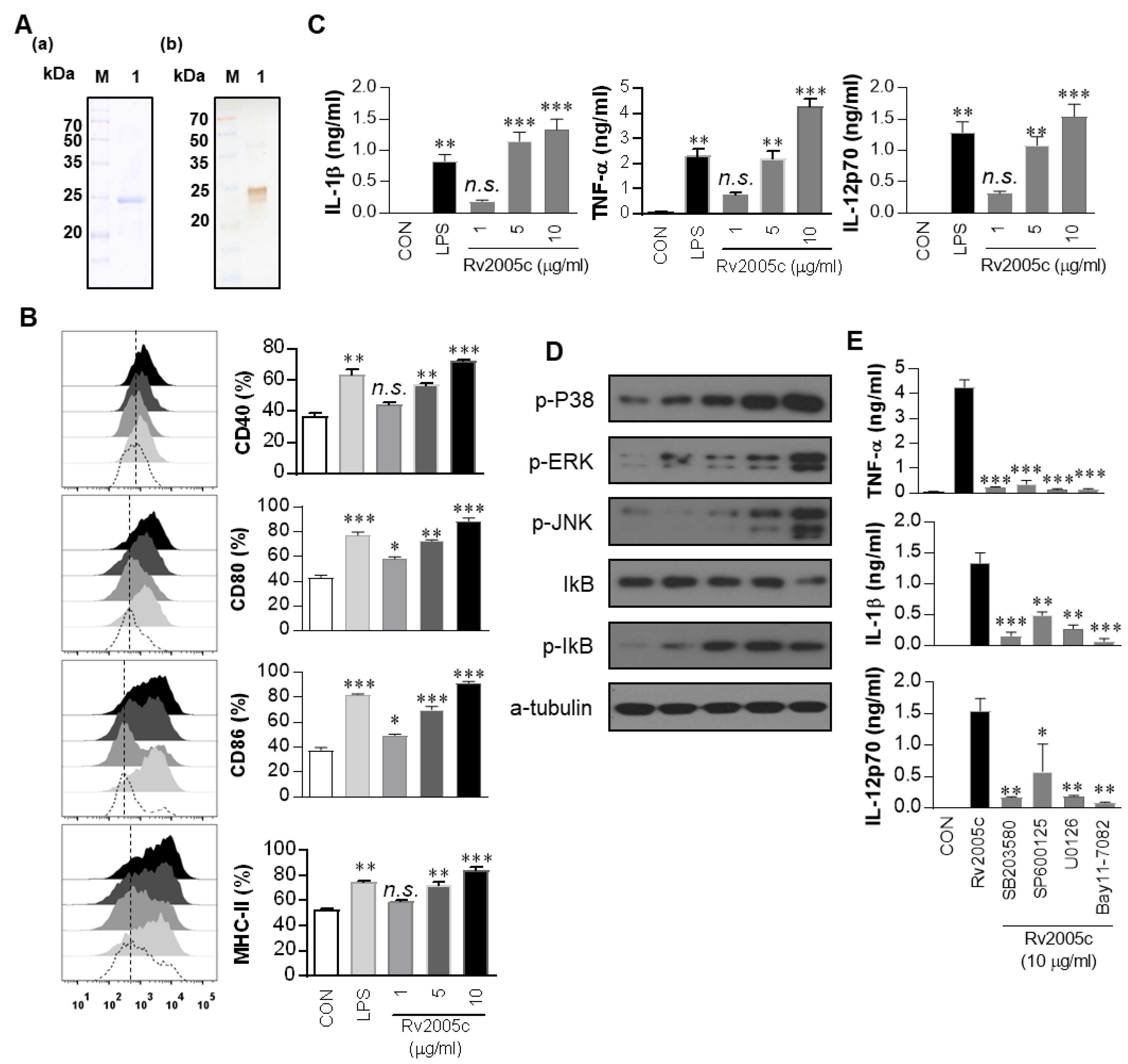
3.1. The Recombinant Rv2005c Protein Induces the Maturation and Activation of Dcs
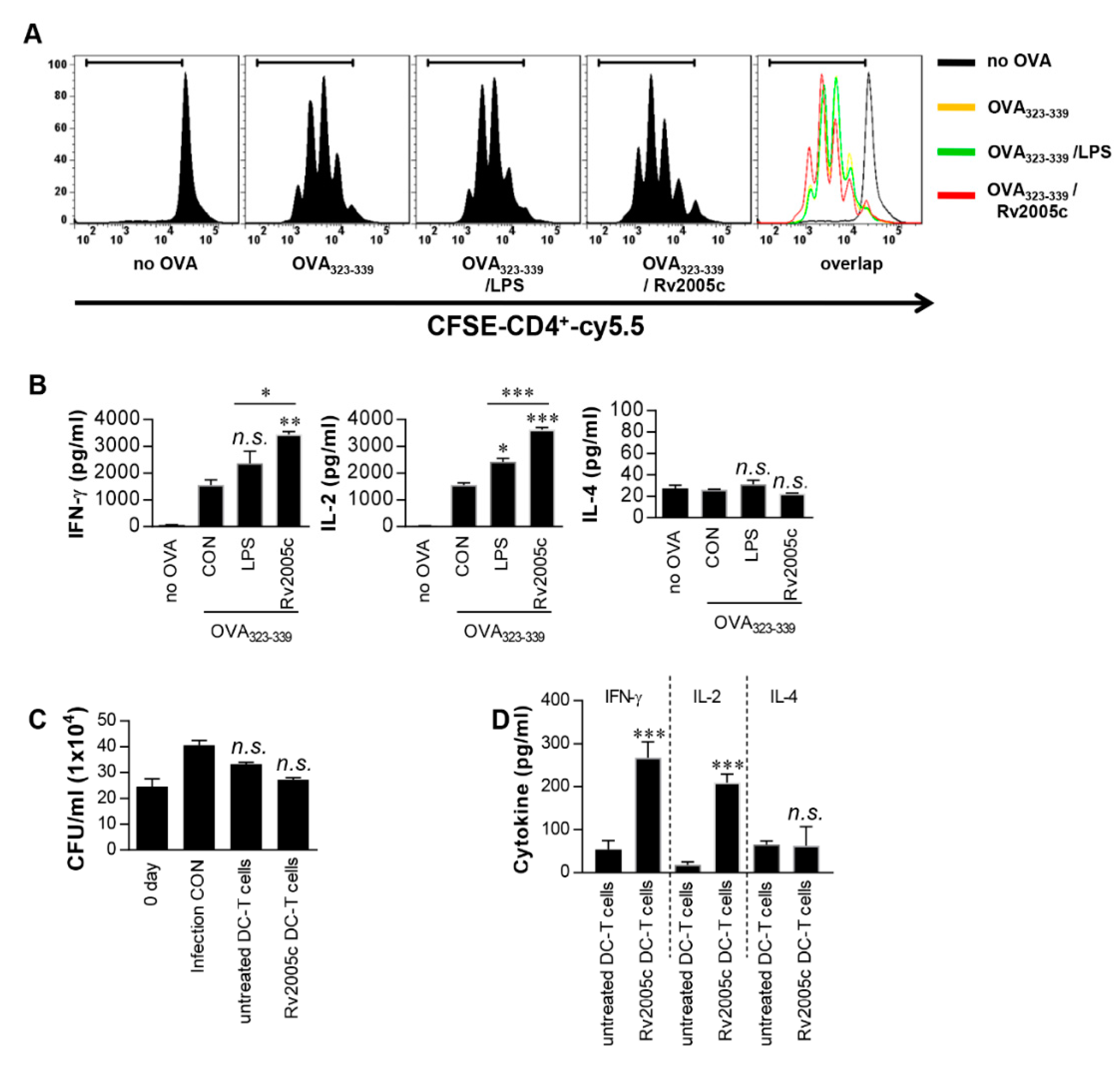
3.2. T Cells Activated by Rv2005c-Matured DCs Do Not Inhibit Intracellular Mtb Growth
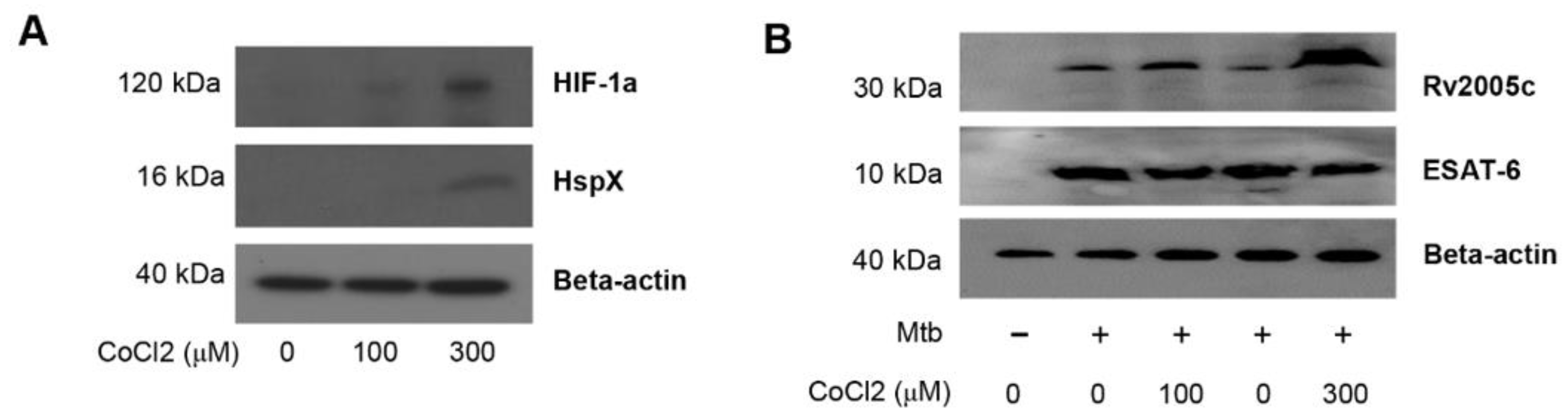
3.3. Increased Expression of Rv2005c by Mtb Within Macrophages under Hypoxic Conditions Induces Intracellular Bacterial Growth Inhibition by T Cells Activated by Rv2005c-Matured DCs
3.4. Rv2005c Does Not Exhibit Significant Vaccine Efficacy
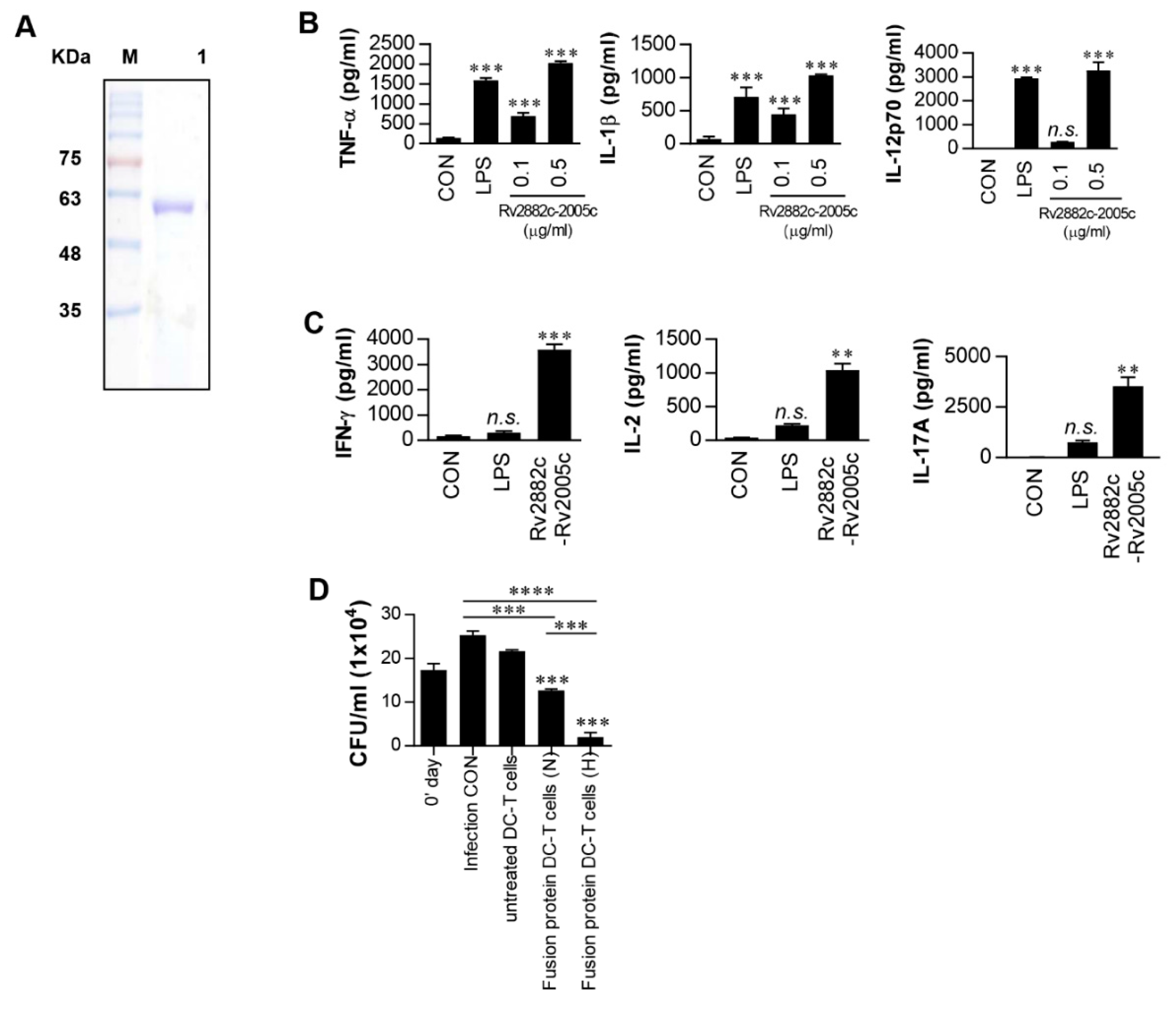
3.5. Intracellular Mtb Growth Inhibition by T Cells Activated by Rv2882c-Rv2005c Fusion Protein-Matured DCs Is Enhanced by Hypoxic Conditions
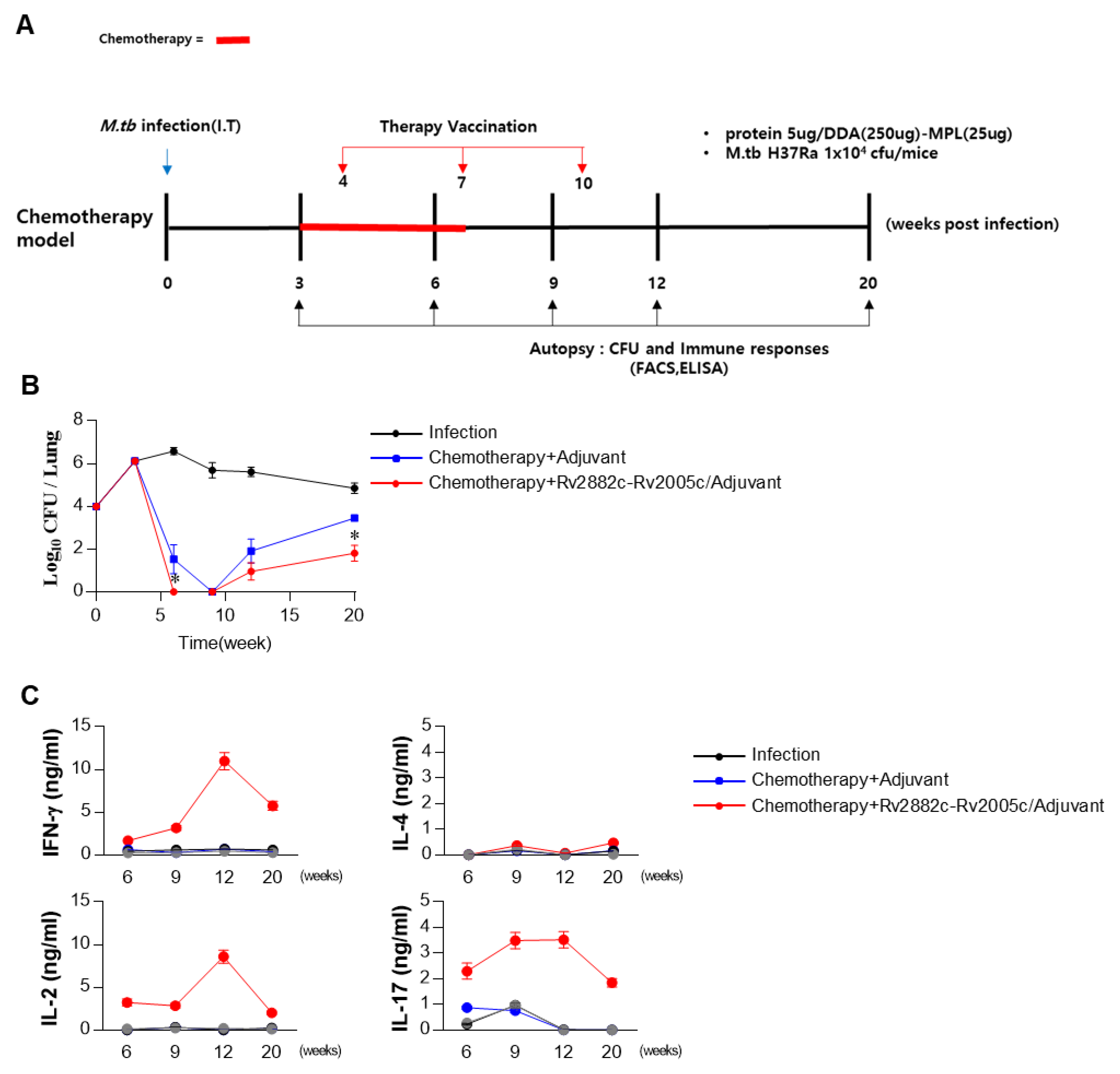
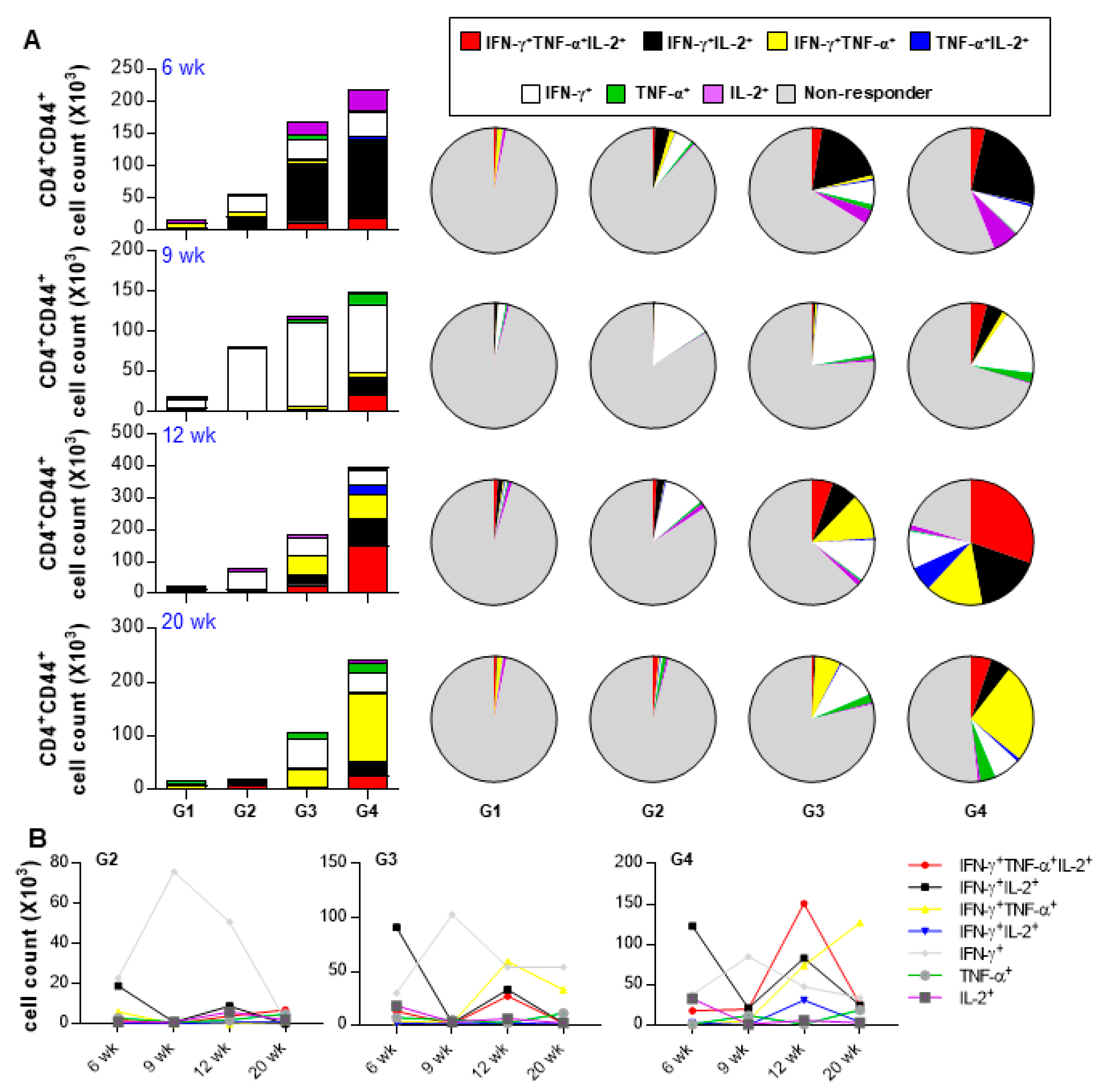
3.6. Rv2882c-Rv2005c Protein Shows Significant Immunotherapeutic Effects
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Bloom, B.R.; Atun, R.; Cohen, T.; Dye, C.; Fraser, H.; Gomez, G.B.; Knight, G.; Murray, M.; Nardell, E.; Rubin, E.; et al. Tuberculosis. In Major Infectious Diseases, 3rd ed.; Holmes, K.K., Bertozzi, S., Bloom, B.R., Jha, P., Eds.; The International Bank for Reconstruction and Development/The World Bank: Washington, DC, USA, 2017. [Google Scholar] [CrossRef]
- Hokey, D.A.; Ginsberg, A. The current state of tuberculosis vaccines. Hum. Vaccin. Immunother. 2013, 9, 2142–2146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seung, K.J.; Keshavjee, S.; Rich, M.L. Multidrug-resistant tuberculosis and extensively drug-resistant tuberculosis. Cold Spring Harb. Perspect. Med. 2015, 5, a017863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schaberg, T.; Rebhan, K.; Lode, H. Risk factors for side-effects of isoniazid, rifampin and pyrazinamide in patients hospitalized for pulmonary tuberculosis. Eur. Respiratory J. 1996, 9, 2026–2030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abate, G.; Hoft, D.F. Immunotherapy for tuberculosis: Future prospects. Immunotargets Ther. 2016, 5, 37–45. [Google Scholar]
- Gill, W.P.; Harik, N.S.; Whiddon, M.R.; Liao, R.P.; Mittler, J.E.; Sherman, D.R. A replication clock for mycobacterium tuberculosis. Nat. Med. 2009, 15, 211–214. [Google Scholar] [CrossRef] [Green Version]
- Roy, E.; Lowrie, D.B.; Jolles, S.R. Current strategies in tb immunotherapy. Curr. Mol. Med. 2007, 7, 373–386. [Google Scholar] [CrossRef] [PubMed]
- Wejse, C.; Gustafson, P.; Nielsen, J.; Gomes, V.F.; Aaby, P.; Andersen, P.L.; Sodemann, M. Tbscore: Signs and symptoms from tuberculosis patients in a low-resource setting have predictive value and may be used to assess clinical course. Scand. J. Infect. Dis. 2008, 40, 111–120. [Google Scholar] [CrossRef]
- Andersen, P. Tuberculosis vaccines—an update. Nat. Rev. Microbiol. 2007, 5, 484–487. [Google Scholar] [CrossRef]
- Baldwin, S.L.; Bertholet, S.; Kahn, M.; Zharkikh, I.; Ireton, G.C.; Vedvick, T.S.; Reed, S.G.; Coler, R.N. Intradermal immunization improves protective efficacy of a novel tb vaccine candidate. Vaccine 2009, 27, 3063–3071. [Google Scholar] [CrossRef] [Green Version]
- Dietrich, J.; Weldingh, K.; Andersen, P. Prospects for a novel vaccine against tuberculosis. Vet. Microbiol. 2006, 112, 163–169. [Google Scholar] [CrossRef] [PubMed]
- Condos, R.; Rom, W.N.; Schluger, N.W. Treatment of multidrug-resistant pulmonary tuberculosis with interferon-gamma via aerosol. Lancet 1997, 349, 1513–1515. [Google Scholar] [CrossRef]
- Yang, X.Y.; Chen, Q.F.; Li, Y.P.; Wu, S.M. Mycobacterium vaccae as adjuvant therapy to anti-tuberculosis chemotherapy in never-treated tuberculosis patients: A meta-analysis. PLoS ONE 2011, 6, e23826. [Google Scholar] [CrossRef] [Green Version]
- Mihret, A. The role of dendritic cells in mycobacterium tuberculosis infection. Virulence 2012, 3, 654–659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakamoto, K. The pathology of mycobacterium tuberculosis infection. Vet. Pathol. 2012, 49, 423–439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dumortier, H.; van Mierlo, G.J.; Egan, D.; van Ewijk, W.; Toes, R.E.; Offringa, R.; Melief, C.J. Antigen presentation by an immature myeloid dendritic cell line does not cause ctl deletion in vivo, but generates cd8+ central memory-like t cells that can be rescued for full effector function. J. Immunol. 2005, 175, 855–863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Etna, M.P.; Giacomini, E.; Severa, M.; Coccia, E.M. Pro- and anti-inflammatory cytokines in tuberculosis: A two-edged sword in tb pathogenesis. Semin. Immunol. 2014, 26, 543–551. [Google Scholar] [CrossRef] [PubMed]
- Aagaard, C.; Hoang, T.; Dietrich, J.; Cardona, P.J.; Izzo, A.; Dolganov, G.; Schoolnik, G.K.; Cassidy, J.P.; Billeskov, R.; Andersen, P. A multistage tuberculosis vaccine that confers efficient protection before and after exposure. Nat. Med. 2011, 17, 189–194. [Google Scholar] [CrossRef]
- Voskuil, M.I.; Visconti, K.C.; Schoolnik, G.K. Mycobacterium tuberculosis gene expression during adaptation to stationary phase and low-oxygen dormancy. Tuberculosis 2004, 84, 218–227. [Google Scholar] [CrossRef]
- Rustad, T.R.; Harrell, M.I.; Liao, R.; Sherman, D.R. The enduring hypoxic response of mycobacterium tuberculosis. PLoS ONE 2008, 3, e1502. [Google Scholar] [CrossRef]
- Betts, J.C.; Lukey, P.T.; Robb, L.C.; McAdam, R.A.; Duncan, K. Evaluation of a nutrient starvation model of mycobacterium tuberculosis persistence by gene and protein expression profiling. Mol. Microbiol. 2002, 43, 717–731. [Google Scholar] [CrossRef]
- Choi, H.G.; Choi, S.; Back, Y.W.; Park, H.S.; Bae, H.S.; Choi, C.H.; Kim, H.J. Mycobacterium tuberculosis rv2882c protein induces activation of macrophages through tlr4 and exhibits vaccine potential. PLoS ONE 2016, 11, e0164458. [Google Scholar] [CrossRef] [PubMed]
- Andersen, P.; Kaufmann, S.H. Novel vaccination strategies against tuberculosis. Cold Spring Harb. Perspect. Med. 2014, 4, a018523. [Google Scholar] [CrossRef] [Green Version]
- Xin, Q.; Niu, H.; Li, Z.; Zhang, G.; Hu, L.; Wang, B.; Li, J.; Yu, H.; Liu, W.; Wang, Y.; et al. Subunit vaccine consisting of multi-stage antigens has high protective efficacy against mycobacterium tuberculosis infection in mice. PLoS ONE 2013, 8, e72745. [Google Scholar] [CrossRef] [PubMed]
- Hingley-Wilson, S.M.; Lougheed, K.E.; Ferguson, K.; Leiva, S.; Williams, H.D. Individual mycobacterium tuberculosis universal stress protein homologues are dispensable in vitro. Tuberculosis 2010, 90, 236–244. [Google Scholar] [CrossRef] [Green Version]
- Sherman, D.R.; Voskuil, M.; Schnappinger, D.; Liao, R.; Harrell, M.I.; Schoolnik, G.K. Regulation of the mycobacterium tuberculosis hypoxic response gene encoding alpha -crystallin. Proc. Natl. Acad. Sci. USA 2001, 98, 7534–7539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peddireddy, V.; Doddam, S.N.; Qureshi, I.A.; Yerra, P.; Ahmed, N. A putative nitroreductase from the dosr regulon of mycobacterium tuberculosis induces pro-inflammatory cytokine expression via tlr2 signaling pathway. Sci. Rep. 2016, 6, 24535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feltcher, M.E.; Gunawardena, H.P.; Zulauf, K.E.; Malik, S.; Griffin, J.E.; Sassetti, C.M.; Chen, X.; Braunstein, M. Label-free quantitative proteomics reveals a role for the mycobacterium tuberculosis seca2 pathway in exporting solute binding proteins and mce transporters to the cell wall. Mol. Cell. Proteomics 2015, 14, 1501–1516. [Google Scholar] [CrossRef] [Green Version]
- Zvi, A.; Ariel, N.; Fulkerson, J.; Sadoff, J.C.; Shafferman, A. Whole genome identification of mycobacterium tuberculosis vaccine candidates by comprehensive data mining and bioinformatic analyses. BMC Med. Genomics 2008, 1, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, H.G.; Choi, S.; Back, Y.W.; Paik, S.; Park, H.S.; Kim, W.S.; Kim, H.; Cha, S.B.; Choi, C.H.; Shin, S.J.; et al. Rv2299c, a novel dendritic cell-activating antigen of mycobacterium tuberculosis, fused-esat-6 subunit vaccine confers improved and durable protection against the hypervirulent strain hn878 in mice. Oncotarget 2017, 8, 19947–19967. [Google Scholar] [CrossRef]
- Choi, S.; Choi, H.G.; Shin, K.W.; Back, Y.W.; Park, H.S.; Lee, J.H.; Kim, H.J. Mycobacterium tuberculosis protein rv3841 activates dendritic cells and contributes to a t helper 1 immune response. J. Immunol. Res. 2018, 2018, 3525302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Back, Y.W.; Choi, S.; Choi, H.G.; Shin, K.W.; Son, Y.J.; Paik, T.H.; Kim, H.J. Cell wall skeleton of mycobacterium bovis bcg enhances the vaccine potential of antigen 85b against tuberculosis by inducing th1 and th17 responses. PLoS ONE 2019, 14, e0213536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Botha, T.; Ryffel, B. Reactivation of latent tuberculosis by an inhibitor of inducible nitric oxide synthase in an aerosol murine model. Immunology 2002, 107, 350–357. [Google Scholar] [CrossRef] [PubMed]
- Shin, A.R.; Shin, S.J.; Lee, K.S.; Eom, S.H.; Lee, S.S.; Lee, B.S.; Lee, J.S.; Cho, S.N.; Kim, H.J. Improved sensitivity of diagnosis of tuberculosis in patients in korea via a cocktail enzyme-linked immunosorbent assay containing the abundantly expressed antigens of the k strain of mycobacterium tuberculosis. Clin. Vaccine Immunol. 2008, 15, 1788–1795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gopinath, V.; Raghunandanan, S.; Gomez, R.L.; Jose, L.; Surendran, A.; Ramachandran, R.; Pushparajan, A.R.; Mundayoor, S.; Jaleel, A.; Kumar, R.A. Profiling the proteome of mycobacterium tuberculosis during dormancy and reactivation. Mol. Cell. Proteomics MCP 2015, 14, 2160–2176. [Google Scholar] [CrossRef] [Green Version]
- Ayrapetov, M.K.; Xu, C.; Sun, Y.; Zhu, K.; Parmar, K.; D’Andrea, A.D.; Price, B.D. Activation of hif1alpha by the prolylhydroxylase inhibitor dimethyoxalyglycine decreases radiosensitivity. PLoS ONE 2011, 6, e26064. [Google Scholar] [CrossRef] [Green Version]
- Rosenkrands, I.; Slayden, R.A.; Crawford, J.; Aagaard, C.; Barry, C.E., 3rd; Andersen, P. Hypoxic response of mycobacterium tuberculosis studied by metabolic labeling and proteome analysis of cellular and extracellular proteins. J. Microbiol. 2002, 184, 3485–3491. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.; Saraav, I.; Sharma, S. Immunogenic potential of latency associated antigens against mycobacterium tuberculosis. Vaccine 2014, 32, 712–716. [Google Scholar] [CrossRef] [PubMed]
- Bertholet, S.; Ireton, G.C.; Ordway, D.J.; Windish, H.P.; Pine, S.O.; Kahn, M.; Phan, T.; Orme, I.M.; Vedvick, T.S.; Baldwin, S.L.; et al. A defined tuberculosis vaccine candidate boosts bcg and protects against multidrug-resistant mycobacterium tuberculosis. Sci. Transl. Med. 2010, 2, 53ra74. [Google Scholar] [CrossRef] [Green Version]
- Park, H.D.; Guinn, K.M.; Harrell, M.I.; Liao, R.; Voskuil, M.I.; Tompa, M.; Schoolnik, G.K.; Sherman, D.R. Rv3133c/dosr is a transcription factor that mediates the hypoxic response of mycobacterium tuberculosis. Mol. Microbiol. 2003, 48, 833–843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heuer, M.; Behlich, A.S.; Lee, J.S.; Ribechini, E.; Jo, E.K.; Lutz, M.B. The 30-kda and 38-kda antigens from mycobacterium tuberculosis induce partial maturation of human dendritic cells shifting cd4(+) t cell responses towards il-4 production. BMC Immunol. 2013, 14, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chu, C.Y.; Jin, Y.T.; Zhang, W.; Yu, J.; Yang, H.P.; Wang, H.Y.; Zhang, Z.J.; Liu, X.P.; Zou, Q. Ca ix is upregulated in cocl2-induced hypoxia and associated with cell invasive potential and a poor prognosis of breast cancer. Int. J. Oncol. 2016, 48, 271–280. [Google Scholar] [CrossRef] [PubMed]
- Beisiegel, M.; Kursar, M.; Koch, M.; Loddenkemper, C.; Kuhlmann, S.; Zedler, U.; Staber, M.; Hurwitz, R.; Kaufmann, S.H. Combination of host susceptibility and virulence of mycobacterium tuberculosis determines dual role of nitric oxide in the protection and control of inflammation. J. Infect. Dis. 2009, 199, 1222–1232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coler, R.N.; Bertholet, S.; Pine, S.O.; Orr, M.T.; Reese, V.; Windish, H.P.; Davis, C.; Kahn, M.; Baldwin, S.L.; Reed, S.G. Therapeutic immunization against mycobacterium tuberculosis is an effective adjunct to antibiotic treatment. J. Infect. Dis. 2013, 207, 1242–1252. [Google Scholar] [CrossRef] [PubMed]
- Lewinsohn, D.A.; Lewinsohn, D.M.; Scriba, T.J. Polyfunctional cd4(+) t cells as targets for tuberculosis vaccination. Front. Immunol. 2017, 8, 1262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoang, T.; Aagaard, C.; Dietrich, J.; Cassidy, J.P.; Dolganov, G.; Schoolnik, G.K.; Lundberg, C.V.; Agger, E.M.; Andersen, P. Esat-6 (esxa) and tb10.4 (esxh) based vaccines for pre- and post-exposure tuberculosis vaccination. PLoS ONE 2013, 8, e80579. [Google Scholar] [CrossRef] [Green Version]
- Chauhan, P.; Jain, R.; Dey, B.; Tyagi, A.K. Adjunctive immunotherapy with alpha-crystallin based DNA vaccination reduces tuberculosis chemotherapy period in chronically infected mice. Sci. Rep. 2013, 3, 1821. [Google Scholar] [CrossRef]







© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Back, Y.W.; Shin, K.W.; Choi, S.; Park, H.-S.; Lee, K.-I.; Choi, H.-G.; Kim, H.-J. Mycobacterium tuberculosis Rv2005c Induces Dendritic Cell Maturation and Th1 Responses and Exhibits Immunotherapeutic Activity by Fusion with the Rv2882c Protein. Vaccines 2020, 8, 370. https://doi.org/10.3390/vaccines8030370
Back YW, Shin KW, Choi S, Park H-S, Lee K-I, Choi H-G, Kim H-J. Mycobacterium tuberculosis Rv2005c Induces Dendritic Cell Maturation and Th1 Responses and Exhibits Immunotherapeutic Activity by Fusion with the Rv2882c Protein. Vaccines. 2020; 8(3):370. https://doi.org/10.3390/vaccines8030370
Chicago/Turabian StyleBack, Yong Woo, Ki Won Shin, Seunga Choi, Hye-Soo Park, Kang-In Lee, Han-Gyu Choi, and Hwa-Jung Kim. 2020. "Mycobacterium tuberculosis Rv2005c Induces Dendritic Cell Maturation and Th1 Responses and Exhibits Immunotherapeutic Activity by Fusion with the Rv2882c Protein" Vaccines 8, no. 3: 370. https://doi.org/10.3390/vaccines8030370