


Abstract
Type 1 diabetes results from the autoimmune destruction of insulin-producing β cells in the pancreas. Genetic and, as yet undefined, environmental factors act together to precipitate the disease. The excess mortality associated with the complications of type 1 diabetes and the increasing incidence of childhood type 1 diabetes emphasize the importance of therapeutic strategies to prevent this chronic disorder. Why is it considered that type 1 diabetes might be preventable? Different strands of diabetes research are coming together to suggest therapeutic targets. Islet cell autoantibody assays make it possible to accurately identify people at risk of future disease. In most cases, a long prodrome provides a window of opportunity to reverse the autoimmune process. Although no current “cure” exists, recent genetic data and preliminary trial results suggest T cells as a target for preventive strategies. Another potentially attainable target is induction of tolerance to the β-cell proteins such as insulin that are inappropriately recognized. Other strategies involve β-cell replacement, but currently there are insufficient donor cells available. This may be overcome as the processes controlling the differentiation of pancreatic and nonpancreatic progenitors as well as replication of existing islet β cells are unravelled.
Type 1 diabetes is characterized by autoimmune destruction of insulin-producing β cells in the pancreas by CD4+ and CD8+ T cells and macrophages infiltrating the islets.1 The disease accounts for about 10% of all cases of diabetes, occurs most commonly in people of European descent and affects 2 million people in Europe and North America. There is a marked geographic variation in incidence, with a child in Finland being about 400 times more likely than a child in Venezuela to acquire the disease (Fig. 1). The current global increase in incidence of 3% per year is well reported,2 and it is predicted that the incidence of type 1 diabetes will be 40% higher in 2010 than in 1998.3 This rapid rise strongly suggests that the action of the environment on susceptibility genes contributes to the evolving epidemiology of type 1 diabetes.

Fig. 1: Geographic variation in annual incidence of type 1 diabetes.
Before safe and rational therapies can be offered in a clinical setting, a detailed understanding of the immune-mediated process that results in type 1 diabetes is required, as is the accurate identification of those at risk of the disease. The immunogenetics of type 1 diabetes has become the model upon which other complex disorders are studied, and in this article we review the importance of recent insights into the pathogenesis and natural history of type 1 diabetes and consider current therapeutic strategies.
Genes: How important are they?
Like other organ-specific autoimmune diseases, type 1 diabetes has human leukocyte antigen (HLA) associations, but how well are they understood? The HLA on chromosome 6 was the first locus shown to be associated with the disease by candidate gene studies4,5 and is considered to contribute about half of the familial basis of type 1 diabetes.6,7 Two combinations of HLA genes (or haplotypes) are of particular importance: DR4-DQ8 and DR3-DQ2 are present in 90% of children with type 1 diabetes.8 A third haplotype, DR15-DQ6, is found in less than 1% of children with type 1 diabetes, compared with more than 20% of the general population, and is considered to be protective.9 The genotype combining the 2 susceptibility haplotypes (DR4-DQ8/DR3-DQ2) contributes the greatest risk of the disease and is most common in children in whom the disease develops very early in life.10 First-degree relatives of these children are themselves at greater risk of type 1 diabetes than are the relatives of children in whom the disease develops later.11
Candidate gene studies also identified the insulin gene on chromosome 11 as the second most important genetic susceptibility factor, contributing 10% of genetic susceptibility to type 1 diabetes.12 Shorter forms of a variable number tandem repeat in the insulin promoter are associated with susceptibility to the disease, whereas longer forms are associated with protection.13 Demonstration of increased expression of insulin (mRNA) in the thymus of people with “long” or protective repeats — which suggests more efficient deletion of insulin-specific T cells during induction of central tolerance — provides an attractive potential mechanism for the role of the insulin gene in type 1 diabetes.14,15
Over the last decade, whole genome screens have indicated that there are at least 15 other loci associated with type 1 diabetes,16–18 and of those, another 2 genes intimately associated with T-cell activation have been identified recently. An allele of the gene for a negative regulator of T-cell activation, cytotoxic T lymphocyte antigen 4 (CTLA-4), found on chromosmone 2q33, is considered to be the third susceptibility locus for type 1 diabetes and has been associated with increased levels of soluble CTLA-419 and the frequency of regulatory T cells.20 A variant of PTPN22, the gene encoding LYP, also a suppressor of T-cell activation, has been deemed the fourth susceptibility factor.21,22 The observation that the 4 most important susceptibility genes for type 1 diabetes can all be represented on a single diagram of antigen presentation to T cells (Fig. 2) emphasizes the potential importance of current therapeutic strategies targeting this interaction. It is also worth noting that the HLA, CTLA-4 and PTPN22 have all been implicated in autoimmune thyroid disease and other autoimmune diseases,23 which supports the premise that similar or overlapping biological pathways contribute to different autoimmune diseases.24
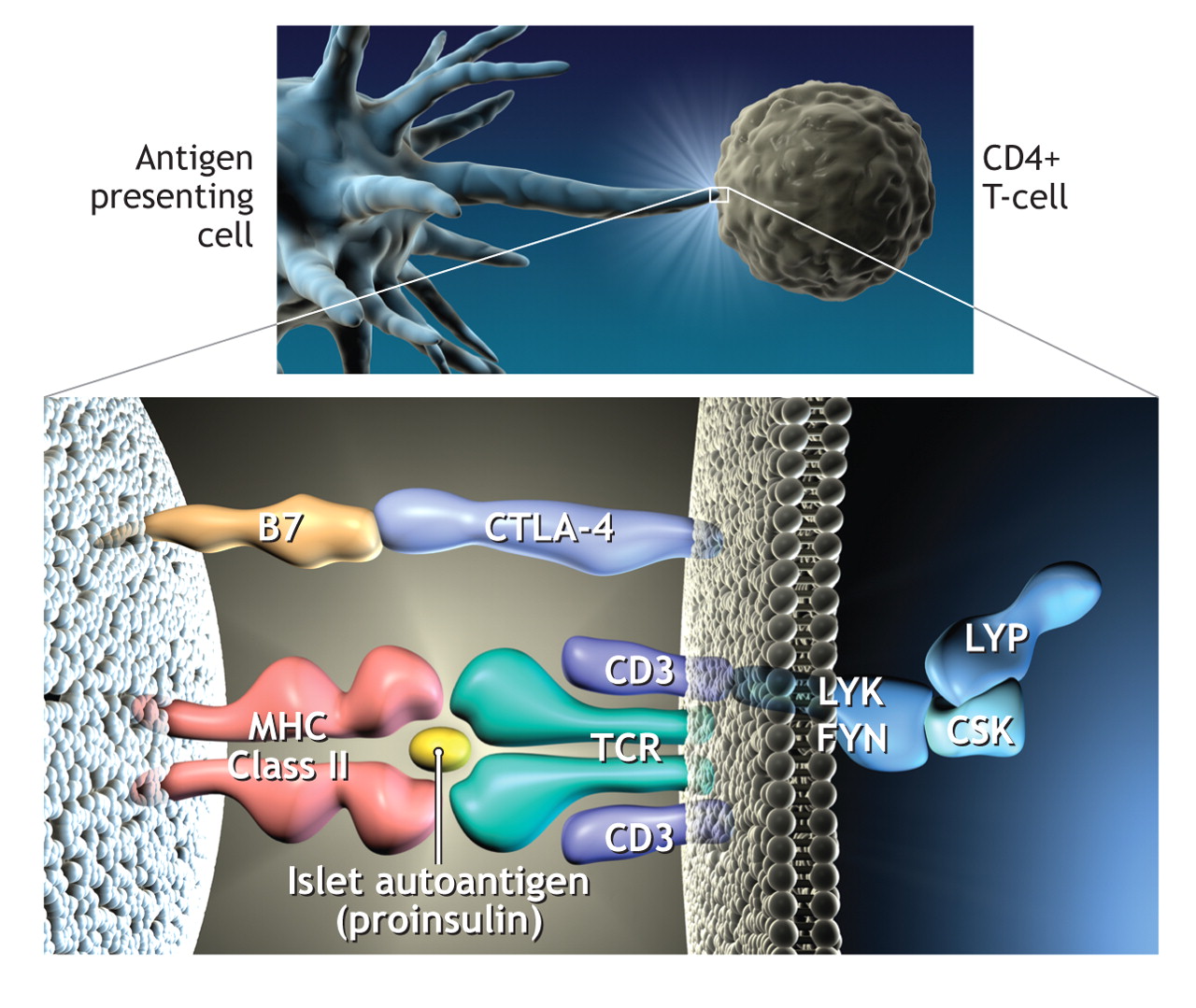
Fig. 2: Representation of the process whereby antigen (in this case peptides of proinsulin) is presented to CD4 T cells by human leukocyte antigen (HLA) class II molecules on the antigen presenting cell. This results in T-cell activation. In this diagram the 4 major genes associated with type 1 diabetes are present. CTLA-4 is an inhibitor of T-cell activation, as is lymphoid tyrosine phosphatase (LYP), which is encoded by the gene PTPN22. The complex of LYP–C-terminal Src kinase (CSK) inhibits Lck signalling after engagement of the T-cell antigen receptor (TCR). Photo by: Lianne Friesen and Nicholas Woolridge
Genetic studies have highlighted the importance of large, well-characterized populations in the identification of susceptibility genes for type 1 diabetes. Recruitment of increasingly large populations of patients with type 1 diabetes and their families is underway (www.t1dgc.org) to provide statistically powerful cohorts in which to identify other disease-associated genes. Some genes will have a relatively minor individual impact on susceptibility to disease but could nevertheless provide more clues to future preventive therapies. The genes for intercellular adhesion molecule (ICAM) and vitamin D are candidates. Some epidemiologic observations support a protective role for vitamin D in type 1 diabetes. Maternal intake of vitamin D in pregnancy and high doses of vitamin D supplements early in life have been shown to protect against islet autoimmunity in offspring,25,26 whereas children with a diagnosis of rickets in the first year of life have been found to have a 3-fold increased risk of type 1 diabetes later in life.27 Further safety studies of vitamin D doses in pregnancy are required.28 Genetic data on type 1 diabetes and vitamin D remain controversial.29,30 The gene for ICAM-1, associated with an increased risk of type 1 diabetes, may contribute to the homing and activation of mononuclear cells in the islets during infection and an early autoimmune response, although confirmatory studies are required.31
Genetic and environmental interactions
Studies in most populations confirm an increase in the incidence of type 1 diabetes, particularly among young children, with the greatest increase occurring in the previously low-incidence countries of eastern Europe. Some studies,32,33 however, have shown convincingly that the increases among young children are occurring because of a shift to lower age at onset rather than an overall increase in incidence in all age groups. These changes are too rapid to be caused by alterations in the genetic background and are likely the result of environmental changes. This is confirmed by recent experiments showing that the increase in type 1 diabetes has been accompanied by a concomitant widening of the HLA risk profile, which suggests increased environmental pressure on susceptible genotypes.34,35
Identification of such environmental factors has proved frustratingly difficult. The most popular candidates are viruses, with enteroviruses,36 rotavirus37 and rubella being suspects. The strongest data to date have supported a role for rubella. Infants infected with congenital rubella syndrome are said to be at increased risk of type 1 diabetes.38 Yet Finland, where vaccination has effectively eradicated rubella, still has one of the highest incidences of type 1 diabetes.39 There is also evidence that some enteroviruses (e.g., Coxsackie B viruses) are less prevalent in countries with high incidences of type 1 diabetes (e.g., Finland) than in countries with low incidences but geographically similar populations (e.g., Russian Karelia).40 This observation may be in keeping with the concept of the hygiene hypothesis,41,42 which proposes that environmental exposure to microbes, other pathogens and their products early in life promotes innate immune responses that suppress atopy and perhaps autoimmunity. In Western cultures, the developing immune system of the infant is no longer exposed to widespread infection, which may contribute to the current increases in incidence observed in atopic and autoimmune disease.
Large studies are required to address the role of environmental factors in susceptibility to type 1 diabetes. An international consortium — the Environmental Determinants of Diabetes in the Young (TEDDY; www.niddk.nih.gov/patient/TEDDY/TEDDY.htm) — has been established to follow several thousand babies with high-risk HLA genotypes from birth until adolescence to identify infectious agents, dietary factors or other environmental factors that trigger islet autoimmunity in genetically susceptible people.
Do we know who is at greatest risk?
More than 30 years ago, it was recognized that antibodies in sera from patients with type 1 diabetes could bind to sections of pancreatic islets. These antibodies were termed islet cell antibodies. The 3 principal autoantigens identified were glutamic acid decarboxylase (GAD 65),43 a protein tyrosine phosphatase-like molecule (IA-2)44 and insulin.45 Studies involving first-degree relatives gave insight into the potential usefulness of islet cell antibodies as predictors of future disease, but the immunoflourescence assay proved difficult to standardize. International workshops to standardize assays for antibodies to GAD, IA-2 and insulin have been more successful,46,47 and it is now clear that about 90% of people with newly diagnosed type 1 diabetes have autoantibodies to at least 1 of these 3 antigens. There is variability in the pattern of humoural immunity, however: insulin autoantibodies are more prevalent in young children,48 IA-2 antibodies often decrease after diagnosis,49 and antibodies to GAD tend to persist.50 It is in the pre-diabetes phase that islet autoantibodies have been most useful. They appear to be already present in most cases of future diabetes by the age of 5 years.51 This indicates that the autoimmune process “smoulders” subclinically for many years in the majority of patients and that clinical symptoms do not appear until up to 80% of β cells have been destroyed.
The observations that islet cell autoantibodies predict autoimmune diabetes in first-degree relatives,52 that the presence of 2 or more autoantibodies in people in the general population is highly predictive of future disease53 and that recent reports have shown that people who have IA-2 antibodies are at very high risk54,55 have opened the way for intervention strategies to delay or slow the autoimmune process. Identification of appropriate agents to reverse or delay the autoimmune process in people found to have 2 or more islet cell autoantibodies is one of the main targets of diabetes research.
Prevention of the disease process before the appearance of islet cell autoantibodies would be ideal, but the accuracy of prediction based on the presence of genes associated with type 1 diabetes is limited. In a study involving patients with islet cell autoantibodies in Belguim, HLA class II genotyping identified a subgroup who represented less than 10% of the Belgian population but who accounted for the majority of future cases of type 1 diabetes in childhood or early adulthood.56 Targeting 10% of the population for therapeutic intervention, however, when the vast majority will not go on to have type 1 diabetes would require a highly effective and safe intervention. Population screening strategies for islet cell autoantibodies can be aided by HLA screening strategies, since 90% of people in whom type I diabetes will later develop are found to be positive for one or both of the HLA susceptibility haplotypes (DR3-DQ2 and DR4-DQ8) and negative for the protective haplotype (DR2-DQ6). This strategy is being used by the type 1 Diabetes Prediction and Prevention Study (DIPP)57 to test the effectiveness of intranasally applied insulin as an antigen-specific therapy.
Therapeutic interventions
Two major trials of ways to prevent or delay the onset of type 1 diabetes are complete.
The European Nicotinamide Diabetes Intervention Trial (ENDIT), a randomized double-blind placebo-controlled trial of high-dose nicotinamide therapy, recruited first-degree relatives of people who were less than 20 years old when their type 1 diabetes was diagnosed, were ICA positive, were less than 40 and had a nondiabetic oral glucose tolerance test. Although nicotinamide had proved protective in animal studies, no effect was observed in the ENDIT study during the 5-year test period.58
A trial that ran concurrently with ENDIT, the Diabetes Prevention Trial – type 1 (DPT-1), studied the efficacy of low-dose insulin injections in high-risk (> 50%) first-degree relatives of patients with type 1 diabetes. In addition, oral insulin capsules were compared with placebo capsules in relatives with a 25%–50% risk of type 1 diabetes. Overall, the insulin treatments had no effect,59 but in a subset of participants in the oral insulin group (those with high levels of insulin autoantibodies), a delay and perhaps a reduction in the incidence of type 1 diabetes was observed.60 Recent data suggesting that insulin is a primary autoantigen in type 1 diabetes61,62 strengthen the case for a therapeutic focus on insulin.
Both the ENDIT and DPT-1 trials, although reporting largely negative findings, have set the standard for future trials and emphasized the requirement for international collaboration to facilitate well-designed trials. To this end, TrialNet (www.diabetestrialnet.org) has been established. TrialNet is a network of 18 clinical centres working in cooperation with screening sites throughout the United States, Canada, Finland, the United Kingdom, Italy, Germany, Australia and New Zealand.
Other studies are examining anti–T-cell strategies. Early studies of cyclosporin in the 1980s provided a proof of principal for the usefulness of immunomodulators in the treatment of type 1 diabetes; the adverse effects of cyclosporin, however, were incompatible with their widespread use.63,64 More sophisticated anti–T-cell strategies have been developed more recently. In one study, hOKT3γ1(Ala-Ala), a humanized, modified anti-CD3 monoclonal antibody, was given to 21 patients within 6 weeks of type 1 diabetes diagnosis; patients in the treatment group had improved C-peptide responses, with effects lasting more than 1 year after a single course of treatment and without chronic immunosuppression.65 In another study, 80 patients with newly diagnosed type 1 diabetes were randomly assigned to receive either an anti-CD3 monoclonal antibody (ChAglyCD3) or placebo; results indicated that residual β-cell function was maintained for at least 18 months and that the effect was strongest in those with the greatest residual β-cell function at study entry.66
In the hOKT3γ1(Ala-Ala) trial, analysis of peripheral blood samples demonstrated an increase in the CD8/CD4 ratio and in particular an increase in CD8+CD25+ regulatory T cells.67 Most studies of regulatory T cells have focused on a subset of naturally occurring CD4+ cells that have the capacity to control self-reactive T cells,68 and their depletion results in autoimmunity.69 Strategies that target the action of regulatory T cells in vivo offer one of the most attractive options for therapy in type 1 diabetes and other autoimmune diseases.
Transplantation
Pancreatic transplantation has offered a successful therapeutic approach for many years.70,71 However, as with all whole-organ transplants, lifelong immunosuppression is required and donor organs are in short supply. An alternative strategy, injection of donor islets into the liver, although less invasive, was found to have mixed success until researchers in Edmonton presented the so-called “Edmonton protocol.”72 With the use of a combination of daclizumab, sirolimus and tacrolimus and islets from more than 1 donor pancreas per recipient, success rates of 80% at 1 year and 20% at 5 years have been reported.73 One of the variables that must be standardized is the crucial islet isolation step.74 Both the quality and the number of islets affect success rates. A prospective multicentre trial coordinated by the Immune Tolerance Network has been established to replicate this success. Like pancreatic transplantation, the procedure is currently limited by availability of donor islets. New sources of functional islets will be required for islet transplantation to make a significant impact on type 1 diabetes.
Regeneration of β cells
The presence of β cells in patients with long-standing type 1 diabetes, despite ongoing autoimmunity, implies that new formation of β cells may be occurring.75 Although an ambitious aim currently, targeted regeneration of such β cells offers another strategy to prevent type 1 diabetes. Regeneration of β cells is therefore an area of major active investigation, with recent studies reporting differentiation of pancreatic and nonpancreatic progenitors as well as replication of existing islet β cells. One study has shown that a single, murine, adult pancreatic precursor exists that can differentiate into cells with the characteristics of islet β cells.76 Another study has shown that pre-existing β cells, rather than pluripotent stem cells, are the main source of new β cells during adult life and after pancreatectomy in mice.77
The future
We are still some way from developing a pill to prevent type 1 diabetes, but all the divergent strands of ongoing research, from epidemiology to molecular biology, immunology to clinical trials, appear to be converging to provide clear perspectives on the therapeutic interventions that are most likely to be successful. Two strategies are open to physicians who have patients with type 1 diabetes: the first is to prevent initiation of autoimmunity; the second is to reverse the effects of ongoing autoimmunity coupled with β-cell regeneration (Fig. 3). Although highly ambitious, the prevention of type 1 diabetes could be possible by identifying and eliminating environmental risk factors. The next line of defence would be to re-educate the immune system through exposure to β-cell antigens with the use of oral or nasal tolerance strategies. The observation that insulin may be the primary autoantigen provides support for therapies using insulin to induce tolerance. The potential to re-educate the immune system, or to divert it using regulatory T cells, and the rapidly expanding field of islet β-cell differentiation give hope that improved strategies to manage this chronic disease are on the horizon.

Fig. 3: Potential targets for therapeutic intervention of type 1 diabetes.
Footnotes
-
This article has been peer reviewed.
Competing interests: None declared.
REFERENCES
- 1.↵
- 2.↵
- 3.↵
- 4.↵
- 5.↵
- 6.↵
- 7.↵
- 8.↵
- 9.↵
- 10.↵
- 11.↵
- 12.↵
- 13.↵
- 14.↵
- 15.↵
- 16.↵
- 17.
- 18.↵
- 19.↵
- 20.↵
- 21.↵
- 22.↵
- 23.↵
- 24.↵
- 25.↵
- 26.↵
- 27.↵
- 28.↵
- 29.↵
- 30.↵
- 31.↵
- 32.↵
- 33.↵
- 34.↵
- 35.↵
- 36.↵
- 37.↵
- 38.↵
- 39.↵
- 40.↵
- 41.↵
- 42.↵
- 43.↵
- 44.↵
- 45.↵
- 46.↵
- 47.↵
- 48.↵
- 49.↵
- 50.↵
- 51.↵
- 52.↵
- 53.↵
- 54.↵
- 55.↵
- 56.↵
- 57.↵
- 58.↵
- 59.↵
- 60.↵
- 61.↵
- 62.↵
- 63.↵
- 64.↵
- 65.↵
- 66.↵
- 67.↵
- 68.↵
- 69.↵
- 70.↵
- 71.↵
- 72.↵
- 73.↵
- 74.↵
- 75.↵
- 76.↵
- 77.↵
In this issue
{kind=link}
{kind=link}
{kind=link}
Article tools






Jump to section
Related Articles
Cited By...
- Network medicine-based epistasis detection in complex diseases: ready for quantum computing
- Structural Changes of Cutaneous Immune Cells in Patients With Type 1 Diabetes and Their Relationship With Diabetic Polyneuropathy
- HLA-DR4Pred2: an improved method for predicting HLA-DRB1*04:01 binders
- Bioinformatics Analysis of next generation sequencing data for Risk Prediction in Patients with Type 1 diabetes mellitus
- Identification of candidate biomarkers and pathways associated with type 1 diabetes mellitus using bioinformatics analysis
- Bromodomain protein inhibition protects {beta}-cells from cytokine-induced death and dysfunction via antagonism of NF-{kappa}B pathway
- Genotyping of coeliac-specific human leucocyte antigen in children with type 1 diabetes: does this screening method make sense?
- Low-Dose Otelixizumab Anti-CD3 Monoclonal Antibody DEFEND-1 Study: Results of the Randomized Phase III Study in Recent-Onset Human Type 1 Diabetes
- Decreased Prevalence of Lymphatic Filariasis Among Subjects with Type-1 Diabetes
- Can Roux-en-Y gastric bypass provide a lifelong solution for diabetes mellitus?
- Streptozocin-Induced Diabetic Mouse Model of Urinary Tract Infection
- Vitamin D supplementation in early childhood and risk of type 1 diabetes: a systematic review and meta-analysis
- The IRF5 polymorphism in type 1 diabetes
- Identification of a WD40 Repeat-Containing Isoform of PHIP as a Novel Regulator of {beta}-Cell Growth and Survival
- Journey to the grave: signaling events regulating removal of apoptotic cells
More in this TOC Section
Similar Articles

