
Abstract
Spinal cord injury (SCI) is a severe condition leading to enduring motor deficits. When lesions are incomplete, promoting spinal cord plasticity might be a useful strategy to elicit functional recovery. Here we investigated whether long-term fluoxetine administration in the drinking water, a treatment recently demonstrated to optimize brain plasticity in several pathological conditions, promotes motor recovery in rats that received a C4 dorsal funiculus crush. We show that fluoxetine administration markedly improved motor functions compared to controls in several behavioral paradigms. The improved functional effects correlated positively with significant sprouting of intact corticospinal fibers and a modulation of the excitation/inhibition balance. Our results suggest a potential application of fluoxetine treatment as a non invasive therapeutic strategy for SCI-associated neuropathologies.
Similar content being viewed by others
Introduction
Spinal cord injury (SCI) is one of the most common causes of disability in adults. This condition affects 2.5 million people in the world, with an incidence of 130,000 new cases every year1. Even if some functional recovery can be observed after incomplete lesions due to spontaneous plasticity-mediated processes, most patients with significant SCI have permanent symptoms and the development of new therapeutic strategies is a major goal of experimental research.
Fluoxetine, a selective serotonin reuptake inhibitor (SSRIs), is a FDA-approved drug widely prescribed in the treatment of various neuropsychiatric disorders. Accumulating evidence suggests that SSRIs can boost neural plasticity in the adult brain, with beneficial effects on learning and memory, hippocampal neurogenesis, synaptogenesis and visual cortex plasticity2,3,4,5. Strikingly, recent studies carried out in animal models and clinical trials highlighted that fluoxetine can induce functional recovery from neurological impairments due to brain injuries and degenerative diseases5,6,7. Indeed, it has been demonstrated that an early prescription of fluoxetine coupled with physiotherapy enhances motor recovery in patients with ischemic stroke8.
Here, we sought to investigate whether boosting neural plasticity with fluoxetine could induce functional recovery in a rat model of spinal cord injury (SCI). We tested the impact of fluoxetine on dorsal funiculus crush at the C4 spinal segment level, a widely studied model of cervical injury leading to partial loss of sensory inputs to the brain and the descending corticospinal control of movements. We analyzed the motor performance of treated and untreated lesioned rats in several behavioral paradigms of forelimb function and fine sensorimotor skills. Our results demonstrated that fluoxetine induced a remarkable recovery of motor behavior compared to untreated animals. Improved motor behavior correlated with specific changes at the level of neuronal sprouting. Moreover, fluoxetine increased the excitation/inhibition ratio in the motor cortex and in the spinal cord of uninjured rats.
Results
In this study we aimed at investigating whether increasing neural plasticity in the rat spinal cord through fluoxetine administration could favor recovery from SCI. In order to maximize the enhancement of neural plasticity operated by fluoxetine, we started our pharmacological treatment three weeks before performing injury, in agreement with a number of studies showing that at least 21 days of treatment are required to induce beneficial effects in the central nervous system9,10.
We administered fluoxetine (FLX, 0.2 g/l in the drinking water) or a normal water drinking regimen (control animals) in adult rats for three weeks. Then, fluoxetine treated animals were subjected to a bilateral crush of dorsal funiculi at the spinal level C4. Control rats were divided in two groups: one group (CTR-inj) received SCI following the same procedure used for FLX animals, the other one (CTR-sham) was only subjected to laminectomy without receiving SCI. The classical kind of lesion used in this study induces a functional damage at the level of both sensory ascending fibers and motor descending dorsal corticospinal tracts (dCST) from the forelimbs11,12, thus affecting the control of fine, skilled movements. After SCI, the pharmacological treatment was not interrupted but continued for one more week in order to maintain fluoxetine levels stable in the acute phase of SCI pathophysiology.
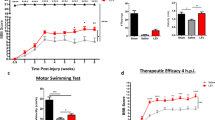
We started to analyze FLX and CTR rat behavior in the Montoya staircase task13, which allows assessing forelimb skilled reaching ability (Fig. 1a, b). We recorded the mean total number of eaten pellets and the performance accuracy (i.e. the ratio between the number of eaten pellets and the sum of eaten and unsuccessfully displaced pellets). Before injury, baseline performance did not differ between FLX and CTR animals (eaten pellets: Mann-Whitney Rank Sum Test, p = 0.420; accuracy: Mann-Whitney Rank Sum Test, p = 0.713). One week after SCI all injured rats were unable to grasp and eat pellets, thus confirming the severity of the lesion received. By the second week post-surgery, FLX animals began to significantly improve their performance compared to CTR-inj rats (Two-Way RM ANOVA, post-hoc Holm-Sidak method, p < 0.05 for both eaten pellets and accuracy). The improvement resulted in a performance of about 60% of pre-injury baseline in the sixth week after SCI, when the CTR-inj group displayed only a 30% of pre-injury performance. The performance of CTR-sham rats did not change throughout the testing period (eaten pellets: One-Way RM ANOVA, p = 0.095; accuracy: One-Way RM ANOVA, p = 0.881).

Fluoxetine induces recovery of motor functions after SCI.
(a, b) Staircase. After SCI, all rats lost grasping ability. Starting from the 2nd week, FLX rats began to retrieve significantly more pellets and showed a greater accuracy than CTR-inj. (c, d) Horizontal ladder. After SCI, the number of footslips was significantly increased in all groups. Starting from the 4th week, FLX rats showed a better performance than CTR-inj. The performance of CTR-sham rats did not change throughout the testing period. Box-whisker plot: the horizontal lines in the box denote the 25th, 50th (median) and 75th percentile values; the small square inside the box represents the mean; error bars denote the 5th and 95th percentile values. X represents max and min values. Curves: error bars represent SEM. Symbols indicate statistical difference.
We further assessed skilled motor control in SCI rats running on a horizontal ladder14 with unevenly spaced rungs (Fig. 1c, d). We recorded the mean number of footslips and the accuracy (i.e. the ratio between the number of correct steps and the sum of total steps) over three consecutive test sessions. Baseline performance was equal in the two groups (footslips, t-test, p = 0.420; accuracy, t-test p = 0.640). However, after injury both FLX and CTR-inj rats showed a dramatic increase in the number of errors made at crossing the ladder. We observed a robust acceleration in the time-course of the recovery process in FLX rats compared to CTR-inj animals (Two-Way RM ANOVA, post-hoc Holm-Sidak method, p < 0.05 for both footslips and accuracy). At the conclusion of the testing period, the performance of FLX animals was indistinguishable from that of CTR-shams (Two-Way RM ANOVA, post-hoc Holm-Sidak method; p = 0.190 for footslips and p = 0.065 for accuracy). The performance of CTR-sham rats did not change throughout the testing period (footslips: One-Way RM ANOVA, p = 0.672; accuracy: One-Way RM ANOVA, p = 0.812).
We also assessed walking patterns with the footprint analysis of gait15,16 (Fig. 2a, b), during which the rat forepaws and hindpaws were stained with different ink colors and the animals were allowed to walk freely over a paper strip. As a sensitive index of walking coordination, we calculated the mean distance between forepaw and hindpaw tracks over at least five consecutive steps. While both injured groups exhibited a reduced coordination in forepaw-hindpaw stepping after SCI, FLX rats showed a marked recovery of gait coordination compared to CTR-inj animals three weeks after SCI (Two Way Repeated ANOVA, post hoc Holm-Sidak method, p < 0.001). The performance of CTR-sham rats did not change throughout the testing period (One-Way RM ANOVA, p = 0.477).

Faster recovery of gait coordination in FLX rats.
(a) Typical footprints of animal walking 2 weeks after SCI. Gray: forepaw footprints; Black: hindpaw footprints (b) Gait analysis. After SCI, FLX rats showed a marked recovery of coordination compared to CTR-inj at the 3rd and 5th testing week. The performance of CTR-sham rats did not change throughout the testing period. Error bars represent SEM. Symbols indicate statistical difference.
In order to investigate possible mechanisms underlying the beneficial effects elicited by fluoxetine, we analyzed sprouting of intact corticospinal fibers (CSTs). After completing the behavioral assessment, we bilaterally injected FLX and CTR-inj rats with the neuronal tracer biotinylated dextran amine (BDA) in the forepaw representation area of the sensorimotor cortex; then, we quantified the number of fibers at increasing distances from the lesion site (Fig 3a). We performed the anatomical analysis in the same animals that were tested in the Montoya staircase task. While no sprouting was observed at any distance in the CTR group, we found that FLX treatment was able to markedly increase sprouting level of CSTs (Two Way ANOVA, p < 0.01, Fig. 3c).

Fluoxetine increases compensatory sprouting after SCI.
(a) Diagram of BDA injections in the sensorimotor cortex. The position of SCI at C4 level is also indicated. (b) Representative pictures of sagittal spinal cord sections at C4 level of CTR-inj (left) and FLX treated (right) rats. Stars indicate lesion cavity. Arrows indicate traced sprouting CST fibers. Scale bar: 50 μm. (c) Quantification of sprouting fibers in the ventral grey matter at 0.5, 1.5, 2.5, 3.5, 4.5 mm from the lesion center, normalized on total counts at medulla oblongata level (100%). FLX rats showed more sprouting fibers at 1.5 and 4.5 mm caudal to the lesion than CTR-inj. Error bars represent SEM. Asterisks indicate statistical significance.
In addition, in order to investigate whether three weeks of fluoxetine treatment were able to promote molecular changes which might favor plasticity in the spinal cord, we analyzed the excitation/inhibition balance in control and fluoxetine treated but not injured animals; the relative strength of the excitatory and inhibitory connections is one major regulator of plasticity in the adult central nervous system17,18 and it is known to be modulated by fluoxetine4,19 (Fig. 4; Supplementary Information, Fig. S1–S2). Quantification of the expression levels of the vesicular transporter proteins for glutamate and GABA (vGluT-1 and vGAT, respectively)20 revealed that the vGluT-1/vGAT expression ratio was significantly increased in fluoxetine treated animals, both in the motor cortex and in the spinal cord (t-test, p < 0.05 in both cases, Fig. 4c). In total agreement with the Western-blot results, we found a significant increase of the Glu/GABA stimulus-evoked overflow ratio in motor cortex and cervical spinal cord synaptosomes of FLX compared to CTR animals (t-test, p < 0.05 and p < 0.01, respectively for motor cortex and spinal cord, Fig. 4d).

Fluoxetine increases the excitation/inhibition ratio in the motor cortex and the spinal cord.
(a) Representative Western blot from control or fluoxetine-treated rat motor cortex (left) and spinal cord (right). Please note that cropped blots are shown (For full length blots, see Supplementary Information, Fig S1–S2). (b) Schematic diagram showing the synaptosome technique. (c, d) Quantification of the expression levels of the vesicular transporter proteins for glutamate and GABA (vGluT-1 and vGAT, respectively) by Western blot analysis and of GABA and glutamate released after a pulse of KCl (15 mM) by synaptosomes in superfusion. Both the vGluT-1/vGAT expression ratio and the Glu/GABA stimulus-evoked overflow ratio were significantly increased after three weeks of fluoxetine treatment, either in the motor cortex and in the spinal cord. Error bars indicate SEM. Asterisks indicate statistical significance.
Discussion
Our findings demonstrate that fluoxetine treatment induces plasticity in the rat spinal cord, creating a permissive condition for functional recovery after the induction of a cervical spinal cord lesion. The establishment of plasticity has been validated by the assessment of the increase of the excitation/inhibition ratio after three weeks of fluoxetine administration. Moreover, the recovery process was accompanied by the sprouting of ventral CST fibers in FLX treated animals, as measured after the end of the behavioral assessment.
In several studies it has been reported that, when SCI is incomplete, processes of partial spontaneous recovery may take place, both in animal models as well as in injured patients and that the functional rescue is accompanied by anatomical reorganizations of neural circuits21. Therefore, development of treatments aimed at increasing plasticity of spinal cord circuits emerges as one of the most promising strategies to restore motor function after injury. In this study, we observed a slight spontaneous recovery of function in untreated controls, in agreement with previous literature22 and a much more pronounced, faster rescue of behavioral performance in FLX rats. The recovery of function induced by fluoxetine administration was associated by the induction of neuronal plasticity, evident at both the anatomical and molecular level. On the one hand, indeed, we observed the presence of sprouting from spared, ventral corticospinal fibers in treated animals, a process which might contribute to restore proper motor functions through functional compensation of the lesioned circuits. One the other hand, the assumption that fluoxetine favors plasticity of spinal cord circuits was reinforced by the finding that the excitation/inhibition ratio was modulated in favor of excitation after three weeks of fluoxetine treatment in non-lesioned rats, both in the motor cortex and in the spinal cord; thus, before the onset of SCI pathophysiology, fluoxetine led to molecular changes that could be permissive for promoting neural plasticity. This is in agreement with previous findings described in studies focusing on the visual cortex4,19 and suggests that a modulation of the relative strength of GABAergic inhibition accompanied with an increase of glutamatergic release might emerge as a critical regulator for plasticity enhancement not only in the adult cerebral cortex but also in the spinal cord.
Even if we focused on the excitatory/inhibitory balance, we can not exclude the possibility that fluoxetine could act on spinal cord plasticity also via other mechanisms. Previous studies demonstrated that chronic antidepressant treatment up-regulates cAMP response element-binding protein (CREB) pathway, which in turns is involved in the increased expression of the neurotrophic factor BDNF23,9. It has been demonstrated that these factors are crucial regulators of synaptic plasticity and neuronal survival; moreover, CREB and BDNF play a pivotal role in neuroplasticity after stroke and SCI24,25,26.
Our results represent a direct proof of principle that the establishment of a more permissive environment through prolonged fluoxetine administration can enhance plasticity in the spinal cord and that this might favor recovery from SCI. A similar link between neural plasticity enhancement and functional improvement after SCI has been previously demonstrated for other treatments, such as chondroitinase ABC12 and anti-NogoA antibody IN-127. In line with these studies, future work should focus on the potentially beneficial effects of a therapeutic treatment with fluoxetine administered after SCI induction. In agreement with this possibility, positive effects of fluoxetine coupled with physiotherapy have been recently shown in patients with ischemic stroke8. Given that fluoxetine requires a prolonged administration period to achieve its functional effects, it is likely that a protocol based on initiating fluoxetine administration immediately after SCI might represent a good strategy to induce recovery.
The non invasive nature of the pharmacological paradigm reported in this study makes fluoxetine administration a treatment with a great potential for clinical application in the field of SCI and related neuropathologies.
Methods
Animal treatment
Adult (2–3 months) male Long Evans rats were used in this study, which was approved by the Italian Ministry of Health (Decreto N° 182/2011-B, 26/09/2011). Three weeks before spinal cord injury (SCI), rats were divided into two groups: control (CTR) and fluoxetine-treated (FLX). Fluoxetine (Fluoxetine-hydrochloride, Galeno, Prato-Italy) was administered in the drinking water (0.2 g/l) as previously described4, corresponding to 16.45 ± 0.36 mg/kg/day. Fluoxetine is known to have a relatively long half life (2 to 4 days and its active metabolite, norfluoxetine, has an extended t1/2 of 7 to 15 days)28, with a wash out of about 3 days in the rat29. The treatment was not interrupted immediately after SCI, but continued until the end of the first postoperative week.
Spinal cord injury
Rats were deeply anesthetized with 1 ml/hg avertin, i.p.. A skin incision (2 cm) was performed in correspondence of the gap between the occipital bone and the dorsal edge of T2 vertebra. Using scissors, the vertebral bone was exposed. C4 dorsal process was lift up with forceps and the dorsal lamina was removed with fine rongeurs. Spinal cord injury was induced inserting the tips of fine forceps 2 mm in depth into the spinal cord parenchyma spanning the gap between the dorsal root entries (1.5 mm lateral to the midline) and down to the spinal canal and keeping them closed for 20 seconds. The injury included the descending dorsal corticospinal tracts (CSTs) and the ascending sensory dorsal columns. All FLX animals were subjected to SCI, while control rats were divided in two groups: one group (CTR-inj) received SCI following the same procedure used for FLX animals, the other one (CTR-sham) was only subjected to laminectomy without receiving SCI.
Behavioral assessment
Montoya staircase reaching task
Before injury, rats were trained for three weeks to remove and eat as many sugar pellets (Bio-serv 45 mg Dustless Precision pellets) as possible from a staircase (Campden Instruments Ltd.)13, until they reached at least 16 pellets eaten on a total of 28 in 15 minutes. After SCI, rats were tested once a week. Two variables were calculated: 1) number of pellets retrieved and eaten; 2) number of pellets displaced on each side of the staircase regardless of whether the animal could grasp and eat them. Accuracy was evaluated as the percentage of displaced pellets successfully eaten. A total of 6 CTR-sham, 13 CTR-inj (injured controls) and 10 FLX rats were included in this experiment.
Horizontal ladder
During the last three days before SCI, animals were trained to walk along an apparatus consisting of side walls made of clear Plexiglas and metal rungs (3 mm diameter), which could be inserted to create an irregular floor with a minimum distance of 1 cm between rungs14. The trials were video recorded over a 60 cm stretch and analyzed offline. After SCI, rats were tested once a week for 6 weeks on an irregular pattern, which was changed for each session. The average number of forepaw steps made over 3 trials per session and the number of footslips was recorded. Accuracy was evaluated as the percentage of correct steps on total. Behavioral testing in the horizontal ladder was performed in a subgroup of the same animals used for the Montoya staircase task (N = 6 CTR-sham, 7 CTR-inj and 7 FLX).
Footprint analysis of gait
Before SCI, the fore and hind paws were painted with dyes of different colors and rats were encouraged to walk in a straight line along a 80 cm long runway over absorbent paper toward home cage15,16. The footprint patterns were then digitalized and analyzed with the Photoshop software to assess coordination. Coordination was measured as the distance between forelimb and hindlimb footprints. A series of at least five sequential steps recorded in the same session was used to determine the mean values of each measurement. Since this task is aimed at assessing the rat spontaneous walking pattern, no training was required before baseline recording. After injury, animals were tested once a week for 6 weeks. In this test, in order to avoid possible confounding effects in gait abilities due to practice in other behavioral task, a separate set of animals were used (N = 6 CTR-sham, 9 CTR-inj and 14 FLX).
Corticospinal tracing and histological assessment
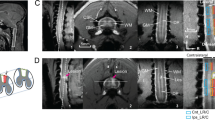
At the end of the functional evaluation, animals were deeply anesthetized with avertin and placed in a stereotaxic frame. Biotinylated dextran amine (10%, BDA-10,000, Invitrogen) was injected stereotaxically using a glass micropipette (Blaubrand®) at a depth of 1.2 mm at six sites distributed over the sensorimotor cortex (stereotaxic coordinates: AP 0, ML ± 2; AP + 1, ML ± 2; AP 2, ML ± 3). After three weeks rats were sacrificed by chloral hydrate overdose and transcardially perfused with PBS followed by 4% paraformaldehyde in 0.1 M phosphate buffer. Medulla oblongata and C3–C5 spinal cord segments were removed and post-fixed in the same solution at 4°C overnight, followed by 30% sucrose. Tissues were frozen in Tissue-Tek® OCT™ (Sakura, The Netherlands) and transverse 50 μm sections were cut with a cryostat. BDA staining was performed with nickel-enhanced diaminobenzidine (DAB) protocol on slides. The accuracy of the lesion was assessed by the quantification of spared tissue above the central canal in sagittal sections of the C3–C5 spinal blocks (Fig. 5). The mean ratio between lesion depth and transverse spinal cord width was used as lesion size index for each animal. Corticospinal sprouting axons were quantified in the gray matter of the C3–C5 spinal blocks. Five vertical (0.5 mm, 1.5 mm, 2.5 mm, 3.5 mm, 4.5 mm) lines were superimposed on each of at least 10 spinal cord sections (Stereoinvestigator) as reference points for crossing axons starting at the center of the lesion. To correct for variability in BDA uptake by corticospinal neurons in the sensorimotor cortex, we normalized the quantitative data by counting BDA-labeled axons in the main pyramidal tract in three transverse sections of medulla oblongata. A total of 7 CTR-inj and 7 FLX rats were included in this experiment.

Spinal cord injury.
(a) Histological sagittal sections of CTR-inj (left) and FLX (right) cervical spinal cord. The star indicates the lesion cavity. (b) Quantification of lesion size in CTR-inj and FLX rats as the percentage of max transverse width of the spinal cord. There is no statistical difference between the two groups. Box-whisker plot: the horizontal lines in the box denote the 25th, 50th (median) and 75th percentile values; the small square inside the box represents the mean; error bars denote the 5th and 95th percentile values. X represents max and min values. Scale bar: 1 mm. Abbreviations: D, dorsal; V, ventral; R, rostral; C, caudal.
Western blot
Uninjured rats (FLX treated for three weeks and CTR) were decapitated and brain and spinal cord were rapidly dissected on ice. The cortical area corresponding to motor cortex and the cervical (C1–C8) portion of the spinal cord were homogenized in an isotonic lysis buffer (50 mM Tris HCl pH 7.6, 0.01% NP40, 150 mM NaCl, 2 mM EDTA, 0.1% SDS, 1 mM PMSF, 0.1 Na3VO4, 1 μg/ml leupeptin, 1 μg/ml aprotinin). Total concentration of samples was assessed with a Bradford assay kit (Bio-Rad, Hercules, CA). Protein extracts were separated using SDS-PAGE (12% Bis-Tris gel, Bio-rad, 1 hr at 200 V), blotted on nitrocellulose membrane, blocked with 4% milk and 0.2% Tween-20 in tris-buffered saline and then probed over night with anti-vGluT1 (1:2,500) or anti-vGAT (1:1,000) rabbit polyclonal antibody (Synaptic Systems, Tubingen, Germany). Blots were then incubated with 1:40,000 horseradish peroxidase (HRP)-conjugated goat anti-rabbit secondary antibody (Jackson Immunoresearch, West Grove, PA). The signal was detected using the luminol/enhancer system (Immun-Star Western C; Bio-Rad) and autoradiography films (HyperFilm; GE Healthcare). As an internal quantification standard, blots were also probed for α-tubulin (monoclonal antibody, 1:10,000, Sigma, Germany). A total of 6 CTR and 6 FLX rats were included in this experiment.
Neurotransmitter quantification
Synaptosome purification
Animals were sacrificed and then the brain and the spinal cord were rapidly removed; motor cortex area and cervical spinal cord (C1–C8) were dissected at 4°C. The tissue was homogenized at 4°C, in 10 volumes of sucrose 0.32 M, buffered with Tris HCl at pH 7.4. The homogenized tissue was centrifuged (5 min, 1000 × g at 4°C and the supernatant was stratified on a four steps discontinuous Percoll® gradient (2, 6, 10, 20% v/v in Tris HCl/sucrose) and again centrifuged (33,500 × g per 5 min a 4°C). After centrifugation, the 10% and 20% Percoll® interface, was collected, washed by centrifugation (15 min, 20,200 × g at 4°C) and then resuspended in a physiologic medium, containing: NaCl 140 mM; KCl 3 mM; MgCl2 1.2 mM; CaCl2 1.2 mM; NaH2PO4 1.2 mM; HEPES 10 mM; glucose 10 mM; pH 7.4. Protein content was measured according to Bradford using bovine serum albumin as a standard. A total of 6 CTR and 6 FLX rats were included in this experiment.
Release experiments
Synaptosomes were incubated at 37°C for 15 min; aliquots of synaptosomal suspension were layered on microporus filters placed at the bottom of a set of parallel superfusion chambers maintained at 37°C (Superfusion System, Ugo Basile, Comerio, Varese, Italy). Superfusion was then started with standard medium at a rate of 0.5 ml/min and continued for 48 min. After equilibration, samples were collected according to the following scheme: two 3-min samples (t = 36–39 min and t = 45–48 min; basal outflow) and one 6-min sample (t = 39–45 min; stimulus-evoked release). A 90-sec period of stimulation was applied at t = 39 min, after the collection of the first sample. Stimulation of synaptosomes was performed with 15 mM KCl, substituting for equimolar concentration of NaCl. Collected samples were analyzed for endogenous glutamate and GABA content. The stimulus-evoked overflow was estimated by subtracting the transmitter content of the two 3-min samples (basal outflow) from the release evoked in the 6-min sample collected during and after the depolarization pulse (stimulus-evoked release). Aminoacid release was calculated as pmol/mg of protein and expressed as Glutamate/GABA ratio.
Neurotransmitter release determination
Endogenous glutamate and GABA content was measured by high performance liquid chromatography analysis following pre-column derivatization with o-phthalaldehyde and gradient separation on a C18 reverse-phase chromatographic column (Chrompack, Middleburg, The Netherlands) coupled with fluorometric detection (excitation wavelength 350 nm; emission wavelength 450 nm;). Homoserine was used as an internal standard.
Statistical analysis
Statistical analysis was performed using Sigma Stat 3.1 (Systat Software, Chicago IL USA).
Multiple groups were compared by ANOVA followed by post-hoc comparisons applying Holm-Sidak test. When two groups were compared, t-test was applied. Normality and homoscedasticity of the data was checked. Data not normally distributed were compared using nonparametric Mann Whitney Rank Sum test. Significance level was equal to 0.05.
Symbols indicate statistical difference: CTR-sham vs CTR-inj, (#) p < 0.05; CTR-sham vs FLX, (+) p < 0.05; FLX vs CTR-inj, (*) p < 0.05.
Ethical statement
Experiments were conducted in conformity with the European Communities Council Directive of November 24, 1986 (86/609/EEC). All experiments were authorized by the Italian Ministry of Health (Decreto N° 182/2011-B, 26/09/2011, Ministero della Salute). Care was taken to minimize the number of animals used and their suffering.
References
Thuret, S., Moon, L. D. F. & Gage, F. H. Therapeutic interventions after spinal cord injury. Nat Rev Neurosci 7, 628–43 (2006).
Malberg, J. E., Eisch, A. J., Nestler, E. J. & Duman, R. S. Chronic antidepressant treatment increases neurogenesis in adult rat hippocampus. J Neurosci 20, 9104–10 (2000).
Hajszan, T., MacLusky, N. J. & Leranth, C. Short-term treatment with the antidepressant fluoxetine triggers pyramidal dendritic spine synapse formation in rat hippocampus. Eur J Neurosci 21, 1299–303 (2005).
Maya Vetencourt, J. F. et al. The antidepressant fluoxetine restores plasticity in the adult visual cortex. Science 320, 385–8 (2008).
Li, W. L., Cai, H. H., Wang, B. & Chen, L. Chronic fluoxetine treatment improves ischemia-induced spatial cognitive deficits through increasing hippocampal neurogenesis after stroke. J Neurosci Res 87, 112–22 (2009).
Narushima, K., Paradiso, S., Moser, D. J., Jorge, R. & Robinson, R. G. Effect of antidepressant therapy on executive function after stroke. Br. J. Psychiatry 190, 260–5 (2007).
Cirrito, J. R. et al. Serotonin signaling is associated with lower amyloid-β levels and plaques in transgenic mice and humans. Proc Natl Acad Sci USA 108, 14968–73 (2011).
Chollet, F., Tardy, J., Albucher, J. F. & Thalamas, C. Fluoxetine for motor recovery after acute ischaemic stroke (FLAME): a randomised placebo-controlled trial. Lancet Neurol 10, 123–30 (2011).
Berton, O. & Nestler, E. J. New approaches to antidepressant drug discovery: beyond monoamines. Nat Rev Neurosci 7, 137–51 (2006).
Baudry, A., Mouillet-Richard, S., Schneider, B., Launay, J. M. & Kellermann, O. miR-16 targets the serotonin transporter: a new facet for adaptive responses to antidepressants. Science 329,1537–41 (2010).
Bradbury, E. J. et al. Chondroitinase ABC promotes functional recovery after spinal cord injury. Nature 416, 636–640 (2002).
García-Alías, G., Barkhuysen, S., Buckle, M. & Fawcett, J. W. Chondroitinase ABC treatment opens a window of opportunity for task-specific rehabilitation. Nat Neurosci 12, 1145–51 (2009).
Montoya, C. P., Campbell-Hope, L. J., Pemberton, K. D. & Dunnett, S. B. The ‘staircase test’: a measure of independent forelimb reaching and grasping abilities in rats. J Neurosci Methods 36, 219–28 (1991).
Metz, G. A. & Whishaw, I. Q. The ladder rung walking task: a scoring system and its practical application. J Vis Exp 28, 10.3791/1204. (2009).
de Medinaceli, L., Freed, W. J. & Wyatt, R. J. An index of the functional condition of rat sciatic nerve based on measurements made from walking tracks. Exp Neurol 77, 634–43 (1982).
Brooks, S. P. & Dunnett, S. B. Tests to assess motor phenotype in mice: a user's guide. Nat Rev Neurosci 10, 519–29 (2009).
Baroncelli, L. et al. Brain plasticity and disease: a matter of inhibition. Neural Plast 2011, 10.1155/2011/286073 (2011).
Bavelier, D., Levi, D. M., Li, R. W., Dan, Y. & Hensch, T. K. Removing Brakes on Adult Brain Plasticity: From Molecular to Behavioral Interventions. J Neurosci 30, 14964–71 (2010).
Chen, J. L. et al. Structural basis for the role of inhibition in facilitating adult brain plasticity. Nat Neurosci 14, 587–94 (2011).
Mainardi, M. et al. Environmental enrichment potentiates thalamocortical transmission and plasticity in the adult rat visual cortex. J Neurosci Res 88, 3048–59 (2010).
Boulenguez, P. & Vinay, L. Strategies to restore motor functions after spinal cord injury. Curr Opin Neurobiol 19, 587–600 (2009).
Weidner, N., Ner, A., Salimi, N. & Tuszynski, M. H. Spontaneous corticospinal axonal plasticity and functional recovery after adult central nervous system injury. Proc Natl Acad Sci USA 98, 3513–8 (2001).
D'Sa, C. & Duman, R. S. Antidepressants and neuroplasticity. Bipolar Disord 4, 183–94 (2002).
Chang, Y. C. et al. Early-life fluoxetine exposure reduced functional deficits after hypoxic-ischemia brain injury in rat pups. Neurobiol Dis 24, 101–13 (2006).
Hannila, S. S. & Filbin, M. T. The role of cyclic AMP signaling in promoting axonal regeneration after spinal cord injury. Exp Neurol 209, 321–32 (2008).
Weishaupt, N., Blesch, A. & Fouad, K. BDNF: the career of a multifaceted neurotrophin in spinal cord injury. Exp Neurol 238, 254–64 (2012).
Merkler, D. et al. Locomotor recovery in spinal cord-injured rats treated with an antibody neutralizing the myelin-associated neurite growth inhibitor Nogo-A. J Neurosci 21, 3665–73 (2001).
Preskorn, S. H. Clinically relevant pharmacology of selective serotonin reuptake inhibitors. An overview with emphasis on pharmacokinetics and effects onoxidative drug metabolism. Clin Pharmacokinet 32, 1–21 (1997).
Caccia, S., Cappi, M., Fracasso, C. & Garattini, S. Influence of dose and route of administration on the kinetics of fluoxetine and its metabolite norfluoxetine in the rat. Psychopharmacology (Berl) 100, 509–14 (1990).
Acknowledgements
We thank Prof. J. Fawcett from University of Cambridge for his precious help and advices on the experimental procedures, Dr. Lucia Galli from the Neuroscience Institute of CNR for her assistance with image acquisition at the microscope and Miss Marilena Marraudino for her help in performing the experiments. Research supported by FIRB (Fondo per gli Investimenti della Ricerca di Base) grant n. RBPR05NWWC.
Author information
Authors and Affiliations
Contributions
L.M., A.S. and M.S. conceived the experiments. M.S., T.B., M.M., M.M., T.B. and G.B. performed the experiments. M.S., A.S. and L.M. wrote the manuscript. All authors discussed the results and commented on the manuscript.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Electronic supplementary material
Supplementary Information
Supplementary information
Rights and permissions
This work is licensed under a Creative Commons Attribution-NonCommercial-ShareALike 3.0 Unported License. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-sa/3.0/
About this article
Cite this article
Scali, M., Begenisic, T., Mainardi, M. et al. Fluoxetine treatment promotes functional recovery in a rat model of cervical spinal cord injury. Sci Rep 3, 2217 (2013). https://doi.org/10.1038/srep02217
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep02217
This article is cited by
-
Buspirone for functional improvement after acute traumatic spinal cord injury: a propensity score-matched cohort study
Spinal Cord (2021)
-
Serotonergic Facilitation of Forelimb Functional Recovery in Rats with Cervical Spinal Cord Injury
Neurotherapeutics (2021)
-
Mapping the physiological and molecular markers of stress and SSRI antidepressant treatment in S100a10 corticostriatal neurons
Molecular Psychiatry (2020)
-
Neutrophil elastase inhibition effectively rescued angiopoietin-1 decrease and inhibits glial scar after spinal cord injury
Acta Neuropathologica Communications (2018)
-
The antidepressant fluoxetine acts on energy balance and leptin sensitivity via BDNF
Scientific Reports (2018)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
