
Key Points
-
Endothelial cells are major participants in and regulators of inflammatory reactions.
-
Resting endothelial cells prevent coagulation, control blood flow and passage of proteins from blood into tissues, and inhibit inflammation. Production of nitric oxide (NO) has a role in these processes and inadequate production of NO is a major cause of endothelial cell dysfunction.
-
Type I activation of endothelial cells mediated by G-protein coupled receptors (GPCRs) that activate G-protein αq subunits and cause endothelial cells to increase blood flow (enhancing delivery of leukocytes to the tissue), increase leakage of plasma proteins into the tissue (creating a provisional matrix to support leukocytes) and promote the binding and activation of neutrophils, encouraging their extravasation into an inflammatory site. Type I activation responses are rapid, independent of protein synthesis, and transient, spontaneously shutting off within 10–20 minutes.
-
Type II activation of endothelial cells mediated by pro-inflammatory cytokines such as tumour-necrosis factor (TNF) and interleukin-1 (IL-1) also increase local blood flow, leakage of plasma proteins and recruit leukocytes. Type II activation responses depend on new gene transcription and protein translation, and are slower in onset but more sustained than type I activation responses, lasting for hours to days.
-
Type-II-activated endothelial cells spontaneously evolve from a phenotype that recruits neutrophils to one that recruits monocytes and T cells. Polarizing cytokines, such as interferon-γ or IL-4 can further modify the activated endothelial cell phenotype to preferentially support T helper 1 (TH1)-cell- or TH2-cell-type inflammatory reactions, respectively.
-
In chronic inflammation, endothelial cells respond to angiogenic factors, such as vascular endothelial growth factor A (VEGFA), to form new blood vessels that are required to sustain an inflammatory neo-tissue such as a pannus in rheumatoid arthritis. Endothelial cells may also respond to lymphotoxin-β to acquire characteristics of high endothelial venules and support the development of tertiary lymphoid organs.
-
Many inflammatory processes display both acute and chronic changes at the same time. This may result because mediators of acute inflammation (such as TNF) contribute to the phenotypes of endothelial cells associated with chronic inflammatory and, similarly mediators of chronic inflammation (such as VEGFA) may also contribute to endothelial cell behaviours associated with acute inflammation.
-
Many therapeutic agents thought to target inflammatory processes or vascular processes affect the inflammatory function of endothelial cells. Our deeper understanding of the mediators, signals and effector molecules involved in endothelial cell inflammatory functions may allow specific targeting of this cell type as a treatment for inflammatory diseases.
Abstract
Inflammation is usually analysed from the perspective of tissue-infiltrating leukocytes. Microvascular endothelial cells at a site of inflammation are both active participants in and regulators of inflammatory processes. The properties of endothelial cells change during the transition from acute to chronic inflammation and during the transition from innate to adaptive immunity. Mediators that act on endothelial cells also act on leukocytes and vice versa. Consequently, many anti-inflammatory therapies influence the behaviour of endothelial cells and vascular therapeutics influence inflammation. This Review describes the functions performed by endothelial cells at each stage of the inflammatory process, emphasizing the principal mediators and signalling pathways involved and the therapeutic implications.
This is a preview of subscription content, access via your institution
Access options
Subscribe to this journal
Receive 12 print issues and online access
$209.00 per year
only $17.42 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout




Similar content being viewed by others
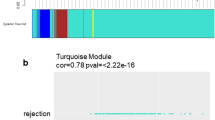
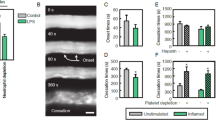
References
Busse, R. & Fleming, I. Vascular endothelium and blood flow. Handb. Exp. Physiol. 176, 43–78 (2006).
Moncada, S. & Higgs, E. A. Nitric oxide and the vascular endothelium. Handb. Exp. Pharmacol. 176, 213–254 (2006).
Arnout, J., Hoylaerts, M. F. & Lijnen, H. R. Haemostasis. Handb. Exp. Physiol. 176, 1–42 (2006).
Minshall, R. D. & Malik, A. B. Transport across the endothelium: regulation of endothelial permeability. Handb. Exp. Pharmacol. 176, 107–144 (2006).
Bazzoni, G. & Dejana, E. Endothelial cell-to-cell junctions: molecular organization and role in vascular homeostasis. Physiol. Rev. 84, 869–901 (2004).
Ley, K. & Reutershan, J. Leucocyte-endothelial interactions in health and disease. Handb. Exp. Pharmacol. 176, 97–133 (2006).
Sessa, W. C. eNOS at a glance. J. Cell Sci. 117, 2427–2429 (2004).
Aird, W. C. Phenotypic heterogeneity of the endothelium: I. Structure, function and mechanisms. Circ. Res. 100, 158–173 (2007).
Gratton, J. P., Bernatchez, P. & Sessa, W. C. Caveolae and caveolins in the cardiovascular system. Circ. Res. 94, 1408–1417 (2004).
Feng, D. et al. Pathways of macromolecular extravasation across microvascular endothelium in resposne to VPF/VEGF and other vasoactive mediators. Microcirculation 6, 23–44 (1999).
Bonfanti, R., Furie, B. C., Furie, B. & Wagner, D. D. PADGEM (GMP140) is a component of Weibel–Palade bodies of human endothelial cells. Blood 73, 1109–1112 (1989).
Middleton, J. et al. Transcytosis and surface presentation of IL-8 by venular endothelial cells. Cell 91, 385–395 (1997).
De Caterina, R. et al. Nitric oxide decreases cytokine-induced endothelial activation. Nitric oxide selectively reduces endothelial expression of adhesion molecules and proinflammatory cytokines. J. Clin. Invest. 96, 60–68 (1995).
Kuhlencordt, P. J. et al. Role of endothelial nitric oxide synthase in endothelial activation: insights from eNOS knockout endothelial cells. Am. J. Physiol., Cell Physiol. 286, C1195–C1202 (2004).
Matsushita, K. et al. Nitric oxide regulates exocytosis by S-nitrosylation of N-ethylmaleimide-sensitive factor. Cell 115, 139–150 (2003).
Hansson, G. K. & Libby, P. The immune response in atherosclerosis: a double-edged sword. Nature Rev. Immunol. 6, 508–519 (2006).
Pober, J. S. & Cotran, R. S. The role of endothelial cells in inflammation. Transplantation 50, 537–544 (1990). This is an extensive review establishing that responses due to endothelial-cell activation can be separated into type I and type II activation responses.
Birch, K. A., Ewenstein, B. M., Golan, D. E. & Pober, J. S. Prolonged peak elevations in cytoplasmic free calcium ions, derived from intracellular stores, correlate with the extent of thrombin-stimulated exocytosis in single human umbilical vein endothelial cells. J. Cell. Physiol. 160, 545–554 (1994).
Luckhoff, A. & Clapham, D. E. Inositol 1,3,4,5-tetrakisphosphate activates an endothelial Ca2+-permeable channel. Nature 355, 356–358 (1992).
Niu, J., Profirovic, J., Pan, H., Vaiskunaite, R. & Voyno-Yasenetskaya, T. G protein βγ subunits stimulate p114RhoGEF, a guanine nucleotide exchange factor for RhoA and Rac1: regulation of cell shape and reactive oxygen species production. Circ. Res. 93, 848–856 (2003).
Egan, K. & FitzGerald, G. A. Eicosanoids and the vascular endothelium. Handb. Exp. Pharmacol. 176, 189–211 (2006).
Mitchell, J. A., Larkin, S. & Williams, T. J. Cyclooxygenase-2: regulation and relevance in inflammation. Biochem. Pharmacol. 50, 1535–1542 (1995).
Stevens, T., Garcia, J. G., Shasby, D. M., Bhattacharya, J. & Malik, A. B. Mechanisms regulating endothelial cell barrier function. Am. J. Physiol., Lung Cell Mol. Physiol. 279, L419–L422 (2000).
Heltianu, C., Simionescu, M. & Simionescu, N. Histamine receptors of the microvascular endothelium revealed in situ with a histamine-ferritin conjugate: characteristic high-affinity binding sites in venules. J. Cell Biol. 93, 357–364 (1982).
Teixeira, M. M., Williams, T. J. & Hellewell, P. G. Role of prostaglandins and nitric oxide in acute inflammatory reactions in guinea-pig skin. Br. J. Pharmacol. 110, 1515–1521 (1993). This is the first demonstration that appreciates the overlapping roles of the endothelial cell mediators the prostaglandins and NO in regulating inflammation-mediated increases in vascular permeability and local blood flow.
Frangogiannis, N. G. Targeting the inflammatory response in healing myocardial infarcts. Curr. Med. Chem. 13, 1877–1893 (2006).
Birch, K. A., Pober, J. S., Zavoico, G. B., Means, A. R. & Ewenstein, B. M. Calcium/calmodulin transduces thrombin-stimulated secretion: studies in intact and minimally permeabilized human umbilical vein endothelial cells. J. Cell Biol. 118, 1501–1510 (1992).
Prescott, S. M., Zimmerman, G. A. & McIntyre, T. M. Human endothelial cells in culture produce platelet-activating factor (1-alkyl-2-acetyl-sn-glycero-3-phosphocholine) when stimulated with thrombin. Proc. Natl Acad. Sci. USA 81, 3534–3538 (1984).
Lorant, D. E. et al. Coexpression of GMP-140 and PAF by endothelium stimulated by histamine or thrombin: a juxtacrine system for adhesion and activation of neutrophils. J. Cell Biol. 115, 223–234 (1991). This first study provides the connection between leukocyte adhesion and activation by juxtacrine signalling.
Marchesi, V. T. The site of leucocyte emigration during inflammation. Q. J. Exp. Physiol. Cogn. Med. Sci. 46, 115–118 (1961).
Schenkel, A. R., Mamdouh, Z. Chen, X, Liebman, R. M. & Muller, W. A. CD99 plays a major role in the migration of monocytes through endothelial junctions. Nature Immunol. 3, 143–150 (2002).
Feng, D., Nagy, J. A., Pyne, K., Dvorak, H. F. & Dvorak, A. M. Neutrophils emigrate from venules by a transendothelial cell pathway in response to FMLP. J. Exp. Med. 187, 903–915 (1998).
Carman, C. V. et al. Transcellular diapedesis is initiated by invasive podosomes. Immunity 26, 784–794 (2007).
Gainetdinov, R. R., Premont, R. T., Bohn, L. M., Lefkowitz, R. J. & Caron, M. G. Desensitization of G protein-coupled receptors and neuronal functions. Annu. Rev. Neurosci. 27, 107–144 (2004).
Pober, J. S. in The Endothelium: A Comprehensive Reference (ed. Aird, W. C.) (Cambrigde Univ. Press, Cambridge, in the press).
Martin, M. U. & Wesche, H. Summary and comparison of the signalling mechanisms of the Toll/interleukin-1 receptor family. Biochim. Biophys. Acta 1592, 265–280 (2002).
Zavoico, G. B., Ewenstein, B. M., Schafer, A. I. & Pober, J. S. IL-1 and related cytokines enhance thrombin-stimulated PGI2 production in cultured endothelial cells without affecting thrombin-stimulated von Willebrand factor secretion or platelet-activating factor biosynthesis. J. Immunol. 142, 3993–3999 (1989).
Pober, J. S. et al. Activation of cultured human endothelial cells by recombinant lymphotoxin: comparison with tumor necrosis factor and interleukin 1 species. J. Immunol. 138, 3319–3324 (1987).
Petrache, I., Birukova, A., Ramierz, S. I., Garcia, J. G. N. & Verin, A. D. The role of microtubules in tumor necrosis factor-α-induced endothelial cell permeability. Am J. Respir. Cell Mol. Biol. 28, 574–581 (2002).
Clark, P. R., Manes, T. D., Pober, J. S. & Kluger, M. S. Increased ICAM-1 expression causes endothelial cell leakiness, cytoskeletal reorganization and junctional alterations. J. Invest. Derm. 127, 762–774 (2006).
Pan, J., Xia, L. & McEver, R. P. Comparison of promoters for the murine and human P-selectin genes suggests species-specific and conserved mechanisms for transcriptional regulation in endothelial cells. J. Biol. Chem. 273, 10058–10067 (1998).
Wang, J. et al. Histamine antagonizes TNF signaling by stimulating TNF receptor shedding from the cell surface and Golgi storage pool. J. Biol. Chem. 278, 21751–21760 (2003).
Karmann, K., Min, W., Fanslow, W. C. & Pober, J. S. Activation and homologous desensitization of human endothelial cells by CD40 ligand, tumor necrosis factor, and interleukin 1. J. Exp. Med. 184, 173–182 (1996).
Munro, J. M., Pober, J. S. & Cotran, R. S. Tumor necrosis factor and interferon-γ induce distinct patterns of endothelial activation and associated leukocyte accumulation in skin of Papio anubis. Am. J. Pathol. 135, 121–133 (1989).
Joris, I., Cuenoud, H. F., Doern, G. V., Underwood, J. M. & Majno, G. Capillary leakage in inflammation. A study by vascular labeling. Am. J. Pathol. 137, 1353–1363 (1990).
Doukas, J. & Pober, J. S. IFN-γ enhances endothelial activation induced by tumor necrosis factor but not IL-1. J. Immunol. 145, 1727–1733 (1990).
Li, J. H. & Pober, J. S. The cathepsin B pathway contributes to TNF plus IFN-γ-mediated human endothelial injury. J. Immunol. 175, 1858–1866 (2005).
Li, J. H. et al. Interferon-γ augments CD95(APO-1/Fas) and pro-caspase-8 expression and sensitizes human vascular endothelial cells to CD95-mediated apoptosis. Am. J. Pathol. 161, 1485–1495 (2002).
Michaeu, O. & Tschopp, J. Induction of TNF receptor I-mediated apoptosis via two sequential signalling complexes. Cell 114, 181–190 (2003).
Kreuz, S., Siegmund, D., Scheurich, P. & Wajant, H. NF-κB inducers upregulate cFLIP, a cycloheximide-sensitive inhibitor of death receptor signaling. Mol. Cell Biol. 21, 3964–3973 (2001).
Bombeli, T., Karsan, A., Tait, J. F. & Harlan, J. M. Apoptotic vascular endothelial cells become procoagulant. Blood 89, 2429–2442 (1997). This study clearly demonstrates that apoptotic endothelial cells become pro-coagulant by increased expression of phosphotidylserine and the loss of anti-coagulant membrane components.
Yoshizumi, M., Perrella, M. A., Burnett, J. C. Jr & Lee, M. E. Tumor necrosis factor downregulates an endothelial nitric oxide synthase mRNA by shortening its half-life. Circ. Res. 73, 205–209 (1993). This study provides the first evidence that pro-inflammatory cytokines, such as TNF, can destabilize endothelial cell NOS as a means to promote inflammation.
Sohn, R. H. et al. Regulation of endothelial thrombomodulin expression by inflammatory cytokines is mediated by activation of nuclear factor-κB. Blood 105, 3910–3917 (2005).
Bevilacqua, M. P. et al. Recombinant tumor necrosis factor induces procoagulant activity in cultured human vascular endothelium: characterization and comparison with the actions of interleukin 1. Proc. Natl Acad. Sci. USA 83, 4533–4537 (1986).
Winsauer, G. & de Martin, R. Resolution of inflammation: intracellular feedback loops in the endothelium. Thromb. Haemost. 97, 364–369 (2007).
Choi, J., Enis, D. R., Koh, K. P., Shiao, S. L. & Pober, J. S. T lymphocyte-endothelial cell interactions. Annu. Rev. Immunol. 22, 683–709 (2004).
Hart, D. N. et al. Localization of HLA-ABC and DR antigens in human kidney. Transplantation 31, 428–433 (1981).
Hancock, W. W., Kraft, N. & Atkins, R. C. The immunohistochemical demonstration of major histocompatibility antigens in the human kidney using monoclonal antibodies. Pathology 14, 409–414 (1982).
Shiao, S. L., McNiff, J. M. & Pober, J. S. Memory T cells and their costimulators in human allograft injury. J. Immunol. 175, 4886–4896 (2005).
Kreisel, D. et al. Vascular endothelium does not activate CD4+ direct allorecognition in graft rejection. J. Immunol. 173, 3027–3034 (2004).
Luster, A. D., Unkeless, J. C. & Ravetch, J. V. γ-interferon transcriptionally regulates an early-response gene containing homology to platelet proteins. Nature 315, 672–676 (1985).
Austrup, F. et al. P- and E-selectin mediate recruitment of T-helper-1 but not T-helper-2 cells into inflammed tissues. Nature 385, 81–83 (1997).
Chaplin, D. D. Cell cooperation in development of eosinophil-predominant inflammation in airways. Immunol. Res. 26, 55–62 (2002).
Briscoe, D. M., Cotran, R. S. & Pober, J. S. Effects of TNF, LPS and IL-4 on the expression of vascular cell adhesion molecule-1 in vivo: Correlation with CD3+ T cell infiltration. J. Immunol. 149, 2954–2960 (1992).
Berg, E. L. et al. The cutaneous lymphocyte antigen is a skin lymphocyte homing receptor for the vascular lectin endothelial cell-leukocyte adhesion molecule 1. J. Exp. Med. 174, 1461–1466 (1991).
Hirahara, K. et al. The majority of human peripheral blood CD4+CD25highFoxp3+ regulatory T cells bear functional skin-homing receptors. J. Immunol. 177, 4488–4494 (2006).
Phillips, G. D., Whitehead, R. A. & Knighton, D. R. Initiation and pattern of angiogenesis in wound healing in the rat. Am. J. Anat. 192, 257–262 (1991).
Paleolog, E. M. Angiogenesis in rheumatoid arthritis. Arthritis Res. 4 (Suppl. 3), 81–90 (2002).
Khurana, R., Simons, M., Martin, J. F. & Zachary, I. C. Role of angiogenesis in cardiovascular disease: a critical appraisal. Circulation 112, 1813–1824 (2005).
Monaco, C., Andreakos, E., Kiriakidis, S., Feldmann, M. & Paleolog, E. T-cell-mediated signalling in immune, inflammatory and angiogenic processes: the cascade of events leading to inflammatory diseases. Curr. Drug Targets Inflamm. Allergy 3, 35–42 (2004).
Shibuya, M. & Claesson-Welsh, L. Signal transduction by VEGF receptors in regulation of angiogenesis and lymphangiogenesis. Exp. Cell Research 312, 549–560 (2006).
Dvorak, H. Vascular permeability factor/vascular endothelial growth factor: a critical cytokine in tumor angiogenesis and a potential target for diagnosis and therapy. J. Clin. Oncol. 20, 4368–4380 (2002).
Qin, L. et al. Down syndrome candidate region 1 isoform 1 mediates angiogenesis through the calcineurin-NFAT pathway. Mol. Cancer Res. 4, 811–820 (2006).
Fiedler, U. & Augustin, H. G. Angiopoietins: a link between angiogenesis and inflammation. Trends Immunol. 27, 552–558 (2006).
Zhang, R. et al. Etk/Bmx transactivates vascular endothelial growth factor 2 and recruits phosphatidylinositol 3-kinase to mediate the tumor necrosis factor-induced angiogenic pathway. J. Biol. Chem. 278, 51267–51276 (2003).
Leahy, K. M., Koki, A. T. & Masferrer, J. L. Role of cyclooxygenases in angiogenesis. Curr. Med. Chem. 7, 1163–1170 (2000).
Strieter, R. M., Burdick, M. D., Gomperts, B. N., Belperio, J. A. & Keane, M. P. CXC chemokines in angiogenesis. Cytokine Growth Factor Res. 16, 593–609 (2005).
Coughlin, C. M. et al. Tumor cell responses to IFNγ affect tumorigenicity and response to IL-12 therapy and antiangiogenesis. Immunity 9, 25–34 (1998).
Drayton, D. L., Liao, S., Mounzer, R. H. & Ruddle, N. H. Lymphoid organ development: from ontogeny to neogenesis. Nature Immunol. 7, 344–353 (2006).
Liao, S. & Ruddle, N. H. Synchrony of high endothelial venules and lymphatic vessels revealed by immunization. J. Immunol. 177, 3369–3379 (2006).
Clauss, M. et al. Vascular permeability factor: a tumor-derived polypeptide that induces endothelial cell and monocyte procoagulant activity, and promotes monocyte migration. J. Exp. Med. 172, 1535–1545 (1990).
Reinders, M. E. et al. Proinflammatory functions of vascular endothelial growth factor in alloimmunity. J. Clin. Invest. 112, 1655–1665 (2003).
Boulday, G., Haskova, Z., Reinders, M. E., Pal, S. & Briscoe, D. M. Vascular endothelial growth factor-induced signalling pathways in endothelial cells that mediate overexpression of the chemokine IFN-γ-inducible protein of 10 kDa in vitro and in vivo. J. Immunol. 176, 3098–3107 (2006).
Lee, C. G. et al. Vascular endothelial growth factor (VEGF) induces remodeling and enhances TH2-mediated sensitization and inflammation in the lung. Nature Med. 10, 1095–1103 (2004).
Fiedler, U. et al. Angiopoietin-2 sensitizes endothelial cells to TNF-α and has a crucial role in the induction of inflammation. Nature Med. 12, 235–239 (2006).
Bhandari, V. et al. Hyperoxia causes angiopoietin 2-mediated acute lung injury and necrotic cell death. Nature Med. 12, 1286–1293 (2006).
Thurston, G., et al. Angiopoietin-1 protects the adult vasculature against plasma leakage. Nature Med. 6, 460–463 (2000).
Kim, I., et al. Angiopoietin-1 negatively regulates expression and activity of tissue factor in endothelial cells. FASEB J. 16, 126–128 (2002).
Madge, L. A. & Pober, J. S. A phosphatidylinositol 3-kinase/Akt pathway, activated by tumor necrosis factor or interleukin-1, inhibits apoptosis but does not activate NFκB in human endothelial cells. J. Biol. Chem. 275, 15458–15465 (2000).
Gullestad, L. & Aukrust, P. Review of trials in chronic heart failure showing broad-spectrum anti-inflammatory approaches. A. J. Card. 95, 17–23 (2005).
Al-Lamki, R. S. et al. TNFR1- and TNFR2-mediated signaling pathways in human kidney are cell type-specific ad differentially contribute to renal injury. FASEB J. 19, 1637–1645 (2005). This reference provides evidence showing that the actions of TNF on endothelial cells may be determined by the pattern of TNFR expression, thereby predicting the outcome for TNF-specific therapy in different disease settings.
Wong, D., Wang, M., Cheng, Y. & FitzGerald, G. A. Cardiovascular hazard and non-steroidal anti-inflammatory drugs. Curr. Opin. Pharmacol. 5, 204–210 (2005).
Sessa, W. C. Can modulation of endothelial nitric oxide synthase explain the vasculoprotective actions of statins. Trends Mol. Med. 7, 189–191 (2001).
Sadeghi, M. M., Collinge, M., Pardi, R. & Bender, J. R. Simvastatin modulates cytokine-mediated endothelial cell adhesion molecule induction: involvement of an inhibitory G protein. J. Immunol. 165, 2712–2718 (2000).
Gilmore, T. D. & Herscovitch, N. Inhibitors of NF-κB signalling: 785 and counting. Oncogene 25, 6887–6899 (2006).
O'Connor, P. Natalizumab and the role of α-4-integrin antagonism in the treatment of multiple sclerosis. Exp. Opin. Biol. Ther. 7, 123–136 (2007).
Weitz-Schmidt, G., Welzenbach, K., Dawson, J. & Kallen, J. Improved lymphocyte function-associated antigen-1 (LFA-1) inhibition by statin derivatives: molecular basis determined by xray analysis and monitoring of LFA-1 conformational changes in vitro and ex vivo. J. Biol. Chem. 279, 46764–46771 (2004).
Ranjbaran, H. et al. Heparin displaces interferon-γ-inducible chemokines (IP-10, I-TAC, and Mig) sequestered in the vasculature and inhibits the transendothelial migration and arterial recruitment of T cells. Circulation 114, 1293–1300 (2006).
Furchgott, R. F. & Zawadzki, J. V. The obligatory role of endothelial cells in the relaxation of arterial smooth muscle by acetylcholine. Nature 288, 373–376 (1980). This reference describes fundamental experiments characterizing an endothelial-cell-derived relaxing factor that led to the discovery of NO.
Acknowledgements
The authors are supported by grants from the National Institutes of Health, USA.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Glossary
- Ectopic lymphoid structures
-
Organized lymphocytic aggregates that form in sites of chronic inflammation. Typically, T-cell- and B-cell-rich zones are segregated, and dendritic cells (DCs), germinal centres with follicular DC (FDC) networks and specialized endothelia are present. These structures are also known as the 'tertiary lymphoid organs' and their formation is termed 'lymphoid neogenesis'.
- Lipid rafts
-
Cholesterol and glycosphingolipid-rich regions of the plasma membrane that provide ordered structure to the lipid bilayer.
- Vesicular–vacuolar organelles
-
A collection of caveolae-like structures, typically located near the inter-endothelial junctions, that may participate in transcytosis of plasma proteins and fluid during certain types of inflammation or in tumour vasculature. VVOs may be distinguished from true caveolae because they are found in caveolin-1 gene deficient mice, which otherwise lack endothelial caveolae.
- Angiogenesis
-
The process of the development of new blood vessels from existing blood vessels. It is frequently associated with tumour development and inflammation. In recent years, it has been appreciated that angiogenesis may be supplemented by the local recruitment of circulating endothelial progenitor cells. The formation of new blood vessels from progenitors is called vasculogenesis.
- E3 ubiquitin ligase
-
An enzyme that is required to attach the molecular tag ubiquitin to proteins. Depending on the position and number of ubiquitin molecules that are attached, the ubiquitin tag can target proteins for degradation in the proteasomal complex, create a scaffold for assembly of signalling complexes, sort them to specific subcellular compartments or modify their biological activity.
- Diapedesis
-
The last step in the leukocyte–endothelial cell adhesion cascade. This cascade includes tethering, triggering, tight adhesion and transmigration. Diapedesis is the migration of leukocytes across the endothelium, which generally occurs by squeezing through the junctions between adjacent endothelial cells, although in some settings, leukocytes have been observed to pass through transiently formed gaps in the cytoplasm of endothelial cells.
- Epitope spreading
-
The de novo activation of autoreactive T cells by self-antigens that have been released after T- or B-cell-mediated bystander damage.
Rights and permissions
About this article
Cite this article
Pober, J., Sessa, W. Evolving functions of endothelial cells in inflammation. Nat Rev Immunol 7, 803–815 (2007). https://doi.org/10.1038/nri2171
Issue Date:
DOI: https://doi.org/10.1038/nri2171
This article is cited by
-
The role of CD146 in renal disease: from experimental nephropathy to clinics
Journal of Molecular Medicine (2024)
-
EASIX and cardiac adverse events after allogeneic hematopoietic cell transplantation
Bone Marrow Transplantation (2024)
-
Proteomic and transcriptomic profiling of brainstem, cerebellum and olfactory tissues in early- and late-phase COVID-19
Nature Neuroscience (2024)
-
The influence of inflammation on the characteristics of adipose-derived mesenchymal stem cells (ADMSCs) and tissue repair capability in a hepatic injury mouse model
Stem Cell Research & Therapy (2023)
-
Posterior reversible encephalopathy syndrome (PRES) associated with SARS-CoV-2 infection in a patient under maintenance haemodialysis: a case report
BMC Nephrology (2023)
