Glymphatic System Dysfunction in Central Nervous System Diseases and Mood Disorders
 Dianjun Zhang
Dianjun Zhang Xinyu Li
Xinyu Li Baoman Li
Baoman Li- 1Department of Forensic Analytical Toxicology, School of Forensic Medicine, China Medical University, Shenyang, China
- 2Liaoning Province Key Laboratory of Forensic Bio-evidence Sciences, School of Forensic Medicine, China Medical University, Shenyang, China
- 3China Medical University Center of Forensic Investigation, School of Forensic Medicine, China Medical University, Shenyang, China
The glymphatic system, a recently discovered macroscopic waste removal system in the brain, has many unknown aspects, especially its driving forces and relationship with sleep, and thus further explorations of the relationship between the glymphatic system and a variety of possible related diseases are urgently needed. Here, we focus on the progress in current research on the role of the glymphatic system in several common central nervous system diseases and mood disorders, discuss the structural and functional abnormalities of the glymphatic system which may occur before or during the pathophysiological progress and the possible underlying mechanisms. We emphasize the relationship between sleep and the glymphatic system under pathological conditions and summarize the common imaging techniques for the glymphatic system currently available. The perfection of the glymphatic system hypothesis and the exploration of the effects of aging and endocrine factors on the central and peripheral regulatory pathways through the glymphatic system still require exploration in the future.
Introduction
The “glymphatic system” (glial-lymphatic system), which was proposed by Jeffrey Iliff, Maiken Nedergaard and her team in 2012, mainly refers to a macroscopic waste clearance system similar to the peripheral lymphoid system in the brain (Iliff et al., 2012). Numerous studies have shown that the glymphatic system exerts a significant effect on the distribution of a variety of substances in the brain, including not only its core waste clearance function but also the distribution and transport of nutrients such as glucose (Lundgaard et al., 2015), some possible treatment substances (recombinant adeno-associated viral vectors) (Murlidharan et al., 2016) and glial calcium signaling (Rangroo Thrane et al., 2013). Because of its important physiological role, the glymphatic system is considered related to the occurrence and development of a variety of diseases (Rasmussen et al., 2018), including neurodegenerative diseases such as Alzheimer’s disease, Parkinson’s disease, as well as mood disorders (Yan et al., 2021).
The brain contains a very rich and complex network of blood vessels. Most of the blood vessels in the brain parenchyma are wrapped by the protruding processes (endfeet) of astrocytes (Nagelhus and Ottersen, 2013; Korogod et al., 2015). The perivascular space (PVS) between the astrocytic endfeet and the vascular wall is filled with cerebrospinal fluid (CSF), which is the main site of action of the glymphatic system. The water channel aquaporin-4 (AQP4) occupies approximately 50% of the surface area of astrocyte processes located close to or in direct contact with blood vessels (Nielsen et al., 1997). CSF passing through the subarachnoid space enters the stroma through AQP4 located in the space around the artery, pushes the interstitial fluid (ISF) to carry solute to the perivenous space, and then flows out of the brain into the cervical lymphatic system (Murtha et al., 2014). Relying on the PVS, astrocytes are the core, and the AQP4-dependent material transport system is the glymphatic system.
The main factors influencing the glymphatic system include AQP4, sleep and arterial pulsation (Jessen et al., 2015). However, the role of arterial pulsation in it is still open to question. AQP4 is a small hydrophobic intrinsic plasma membrane protein, and its physiological function is to promote the transport of water through the plasma membrane. Because its expression on astrocytes is highly polarized (mainly concentrated on the capillary surface of the endfeet) (Nielsen et al., 1997), it reduces the resistance of CSF transport between the PVS and interstitial space (Iliff et al., 2012). Therefore, astrocytes have the ability to guide the transformation of CSF and ISF into each other in both the space around the arteries and veins.
Some studies have shown that the glymphatic system is inhibited in the awake state but enhanced in the sleep state (Xie et al., 2013). Sleep may affect the glymphatic system by changing the volume of the brain stroma. The brain interstitial volume fraction in asleep and anesthetized mice was higher than that in the awake state, which reduced the resistance of CSF flowing into the brain stroma and was beneficial to the operation of the glymphatic system (Xie et al., 2013). In addition, norepinephrine is a major neurotransmitter involved in the awakening state (Berridge and Waterhouse, 2003), and a sudden increase in its levels during awakening increases the cell volume in the brain parenchyma and may resist the inflow of CSF by reducing the interstitial volume (O’Donnell et al., 2012). Although controversy persists regarding whether the choroid plexus is the source of CSF production (Oresković and Klarica, 2010; Trillo-Contreras et al., 2019), several studies have confirmed that norepinephrine directly acts on choroid plexus epithelial cells to inhibit CSF production (Nilsson et al., 1992), which may be part of the mechanism of the sleep inhibitory glymphatic system.
Generally, intracranial arterial pulsation is presumed to be the main physical force promoting the operation of the glymphatic system (Jessen et al., 2015). Related experiments have indicated that the efficiency of CSF flowing into the brain is positively correlated with the pulsation of arteries (cortical penetrating arteries) (Iliff et al., 2013b). Every 10-beat reduction in heart rate reduces waste clearance and leads to an approximately 20% increase in β-amyloid protein (Aβ) levels in the brain parenchyma, while an increase in heart rate of 30 beats per minute produced an approximately 30% decrease in Aβ levels in the brain parenchyma (Kyrtsos and Baras, 2015). Although the glymphatic system is most active during sleep (Xie et al., 2013), blood pressure, cerebral blood flow (Kotajima et al., 2005) and vascular movement (Zhang and Khatami, 2014) are all at a low level, which contradicts this conclusion. However, a study by Goulay et al. (2018) in non-human primates confirmed that solute clearance in the brain is severely impaired in the presence of CSF leakage, supporting a link between CSF circulation and glymphatic system. Therefore, the relationship between arterial pulsation and the glymphatic system is not fully understood.
This review summarizes and introduces the research progress on understanding the role of the glymphatic system in central nervous system diseases and mood disorders, discusses the possible mechanisms underlying the occurrence and development of related diseases. We focus on the abnormalities in various components of the glymphatic system and the role of sleep in the development of the disease, combined with summarizing the commonly used visual imaging methods, to provide strategies for the prevention and treatment of related diseases in the future.
Central Nervous System Diseases
Neurodegenerative Diseases
Neurodegenerative diseases (NDDs) are diseases characterized by progressive dysfunction and neuronal loss (Kovacs, 2016). Common NDDs include Alzheimer’s disease (AD), Parkinson’s disease (PD), amyotrophic lateral sclerosis (ALS) and Huntington’s disease (HD). A possible cause of these destructive diseases is the abnormal aggregation and deposition of toxic proteins such as Aβ, α-synuclein, TAR-DNA-binding protein 43 and mutant huntingtin protein respectively (Kovacs, 2016; Ruz et al., 2020). This pathological deposition occurs not only in the brain but also in peripheral organs (Kovacs, 2019) and affects neurons and glial cells simultaneously (Kovacs, 2016).
Alzheimer’s Disease
Alzheimer’s disease is a relatively slowly progressing neurodegenerative disease. As a common type of dementia, its main pathological features are the deposition of senile plaques and neurofibrillary tangles (NFTs) (De-Paula et al., 2012; Tarasoff-Conway et al., 2015). Senile plaques (also known as neuritic plaques) are attributed to the extracellular deposition of Aβ, and the accumulation of high-density plaques in the hippocampus, amygdala and cerebral cortex leads to the activation of astrocytes and microglia, synaptic loss, and cognitive impairment (Tabaton and Piccini, 2005; Armstrong, 2009; Chen et al., 2017). Aβ deposition also ossifies blood vessels and reduces arterial pulsation, leading to cerebral amyloid angiopathy (CAA) (Serrano-Pozo et al., 2011). Tau is a splice variant of microtubule-associated protein tau (MAPT), which is an intracellular neuronal protein that stabilizes axons (Shea and Beermann, 1994). In patients with AD, intracellular tau (i-tau) is hyperphosphorylated, resulting in the formation of insoluble neurofibrillary tangles that are difficult to remove (Binder et al., 2005).
Schubert et al. (2019) uses MRI and dynamic 11C-PiB PET to prove that CSF clearance deficits in AD associated with Aβ deposits. A study using ultrafast 10 Hz magnetic resonance encephalography also showed that pervasive abnormality in (peri) vascular cerebrospinal fluid impulse propagation exists in patients with AD, which can damage glymphatic brain clearance of Aβ in AD (Rajna et al., 2021). The glymphatic system is inhibited in an AD (APP/PS1) mouse model, and this inhibition occurs with or without showing extensive Aβ deposits, due to the accumulation of toxic Aβ species, such as soluble oligomers (Peng et al., 2016), this indicates that there may be a complex interaction relationship between AD and glymphatic system. Using transgenic mice, Patel et al. (2019) confirmed that the glymphatic system plays an important role in the transport and clearance of extracellular tau protein. As the functional core of the glymphatic system, AQP4 promotes the clearance of the interstitial aggregation of proteins, including Aβ and tau (Mader and Brimberg, 2019). Two-photon imaging studies showed that the Aβ clearance rate of AQP4 knockout mice is decreased by 55–65%(Iliff et al., 2012), indicating that AQP4 plays an important role in the clearance of extracellular Aβ (EAβ) in mice. The pharmacological inhibition of AQP4 (AQP4 inhibitor TGN-020) also significantly reduces the tau clearance rate (Harrison et al., 2020). Using human transcriptomic data, researchers demonstrated that the expression of perivascular astroglial gene products are associated with phosphorylated tau levels in temporal cortex (Simon et al., 2018). However, under the pathological condition of AD, AQP4 may be abnormal. The lower concentrations of Aβ(1-42) (a soluble monomeric isomer of Aβ; Zheng et al., 2012) increased AQP4 expression in cultured mouse cortical astrocytes, while the higher concentrations of Aβ(1-42) decreased AQP4 expression (Yang et al., 2012). AQP4 depolarization was observed in the brains of patients with AD (Zeppenfeld et al., 2017) and AD model mice (Xu et al., 2015). These deletions or changes in the location of polarization substantially inhibit the clearance function of the glymphatic system, resulting in the accumulation of large amounts of Aβ and tau in the brain, leading to the continuous deterioration of AD.
One of the characteristics of patients with AD is sleep disturbance (Moran et al., 2005). Sleep-wake cycle disorders caused by Aβ accumulation and loss of diurnal fluctuations in Aβ levels in ISF are observed in mice and humans (Roh et al., 2012), while the glymphatic system is most active during sleep. The Aβ clearance rate in the brain of sleeping mice is twice as fast as that of awake mice (Xie et al., 2013), and Aβ deposition in the human brain increases significantly after sleep deprivation for one night (Shokri-Kojori et al., 2018). Based on these results, sleep plays an important role in Aβ clearance. Another study showed that people with poor sleep habits and specific genetic variations in AQP4 have more Aβ deposits in the brain (Rainey-Smith et al., 2018), suggesting that AD may inhibit glymphatic system function through both AQP4 abnormalities and sleep disorders, resulting in protein accumulation and related pathological changes.
Parkinson’s Disease
Parkinson’s disease is a common neurodegenerative disease, and its symptoms mainly include motor disorders (neuromuscular stiffness, motor retardation, and tremor) and non-motor disorders (sleep disorders, constipation, and malnutrition) (Henderson et al., 2019). The main pathological feature of PD is the loss of dopaminergic neurons in the substantia nigra, accompanied by the accumulation of α-synuclein (α-syn) in Lewy bodies (Bohnen and Hu, 2019). Blocking meningeal lymphatic drainage by ligating the deep cervical lymph nodes in mice has also been shown to aggravate the pathological and motor symptoms caused by α-syn (Zou et al., 2019), indicating a possible relationship between the glymphatic system and PD.
α-Syn is considered a protein that regulates synaptic neurotransmitter release and autophagy (Bobela et al., 2015), and its deposition in the brain is the basis of the neuropathology of PD. α-Syn accumulates in many parts of the brain, causing neuroinflammation. The interaction between neuroinflammation and systemic oxidative stress may exacerbate reactive astrogliosis and potential damage to the glymphatic system (Verkhratsky et al., 2015; Chiang et al., 2018), while the neurotoxicity of α-syn itself promotes the loss of dopaminergic neurons in the substantia nigra-striatum circuit, leading to related symptoms (Taylor et al., 2002; Kovacs et al., 2008). Some experiments have shown a negative correlation between α-syn deposition and AQP4 expression (Hoshi et al., 2017), while a decrease in AQP4 expression aggravates the degeneration of dopaminergic neurons (Zhang et al., 2016). To some extent, this finding explains the possible pathological relationship between the glymphatic system and PD.
Sleep disorders and poor sleep quality are strongly associated with decreased cognitive performance in patients with PD (Stavitsky et al., 2012; Bohnen and Hu, 2019). One of the common symptoms is rapid eye movement-sleep behavior disorder (RBD), a sleep disorder characterized by a lack of muscle tone, which leads to abnormal body movement during REM sleep and often occurs before motor symptoms (Barber and Dashtipour, 2012). A study of autopsy samples showed a cumulative increase in α-syn levels in many regions of the brains of patients with PD presenting sleep disorders (Kalaitzakis et al., 2013), and evidence that sleep dysfunction in patients with PD is caused by the degeneration of dopaminergic neurons in a pathway similar to RBD has been reported (Dickson et al., 2009; Kalaitzakis and Pearce, 2009; Grinberg et al., 2010). Dopamine, similar to norepinephrine, increases alertness and promotes arousal, and its levels increase mainly in the state of awakening (Isaac and Berridge, 2003; Murillo-Rodríguez et al., 2009). There is a possible interaction between dopamine and clock gene expression, and dopaminergic dysfunction may lead to circadian rhythm disorder (Imbesi et al., 2009; Albrecht, 2013). In addition, some experiments have shown that dopamine reduces the expression of AQP4 in striatal astrocytes, and this downregulation causes the dopamine-induced decrease in proliferation of these cells (Küppers et al., 2008). Based on a large number of experimental results, researchers speculated a vicious cycle, which is the combined neurotoxic effects of α-syn and dopaminergic neurons lead to glymphatic system dysfunction by causing REM sleep abnormalities, circadian rhythm and clock gene dysfunction, while the decrease of α-syn clearance furthers the PD degenerative process (Sundaram et al., 2019).
Amyotrophic Lateral Sclerosis
Amyotrophic lateral sclerosis (ALS) is a rapidly progressive neurodegenerative disease characterized by the death of supraspinal and supraspinal neurons responsible for controlling various muscles (Kiernan et al., 2011). Many proteins are related to ALS, such as superoxide dismutase 1 (SOD1), fusion sarcoma (FUS) and Aaxin-2, among which TAR-DNA-binding protein 43 (TDP-43) is the most relevant; it binds RNA and regulates mRNAs involved in neuronal development (Geuens et al., 2016; Prasad et al., 2019). TDP-43 inclusion bodies are widely regarded as pathological markers of ALS, and more than 95% of patients with ALS show TDP-43 deposition (Neumann et al., 2006). TDP-43 deposition represents a possible glymphatic disorder, and the observation of AQP4 overexpression (Nicaise et al., 2009; Dai et al., 2017) and depolarization (Dai et al., 2017) in mouse or rat ALS models supports this speculation. AQP4 depolarization is consistent with the progression of ALS, and its persistence will interfere with neuronal function (Dai et al., 2017). A recent study observed that the cervical spinal cord clearance of the fluorescent dye-labeled tracer (ovalbumin) injected through the cisterna magna of ALS model SOD1G93A mice decreased by approximately 35% compared with wild-type mice, suggesting that ALS model mice have a dysfunctional glymphatic system (Hirose et al., 2021). Meng et al. (2019) temporarily increase the permeability of the blood-brain barrier, recording gadobutrol patterns in the PVS, subarachnoid space, and space surrounding large cortical draining veins in ALS patients, suggesting glymphatic efflux persists in humans. Another pathological feature of ALS is reactive glial hyperplasia characterized by astrocyte hypertrophy and microglial proliferation (Ng et al., 2015), which may also indicate potential glymphatic system damage.
In addition, sleep quality and respiratory function are negatively correlated with the severity of ALS (Ahmed et al., 2016). Increased norepinephrine concentrations have been detected in the CSF, plasma and spinal cord tissues (thoracic and lumbar spinal cord) of patients with ALS (Bertel et al., 1991), which may also affect the glymphatic system.
Traumatic Brain Injury
Traumatic brain injury (TBI) increases the risk of other neurological and mental disorders, including chronic traumatic encephalopathy, AD, anxiety, depression and personality disorders (Bolte and Lukens, 2021). TBI induces the release of Aβ and C-tau. C-tau is a biomarker of brain injury, and its release is related to the severity of TBI (Gabbita et al., 2005). Several methods (AQP4 gene knockout, cisterna magna cisternostomy, acetazolamide treatment, and sleep deprivation) have been used to inhibit the function of the glymphatic system in the mouse brain. The plasma levels of three biomarkers of TBI [S100β, glial fibrillary acidic protein (GFAP) and neuron-specific enolase] are decreased, indicating that these biomarkers leave the brain through the glymphatic system (Plog et al., 2015), while this glymphatic system-dependent process slows down after TBI (Iliff et al., 2014). In patients with TBI, astroglial inflammation, C-tau, p-tau and Aβ protein waste accumulation and other pathological manifestations are considered to lead to glymphatic system dysfunction (Jessen et al., 2015), and subarachnoid hemorrhage caused by head trauma is also considered to damage the glymphatic system (Gaberel et al., 2014). Inglese et al. (2005) used MRI to evaluate the structural abnormalities of the brain of 24 patients with mild TBI, compared with healthy control subjects, the number of dilated Virchow-Robin spaces (VRS) in the brain of patients with mild TBI increased, indicating the potential dysfunction of glymphatic system. Experiments have suggested that TBI can lead to the clearance of radioactive tracers from the mouse brain: the function of the glymphatic system is reduced by approximately 60% and lasts for at least 28 days (Iliff et al., 2014). This dysfunction of the glymphatic system is related to the depolarization of AQP4 (Ren et al., 2013; Mestre et al., 2018) and has a certain regional specificity (Christensen et al., 2020). As the severity of TBI increases, the recovery time of the AQP4 polarization distribution is correspondingly prolonged (no recovery is observed after 28 days) (Ren et al., 2013). TBI is also related to the formation of astrocyte scars and the continuous activation of neuroinflammation (Iliff et al., 2014). Inflammation and proliferation of reactive astrocytes and microglia appear around the injured site (Giza and Hovda, 2014), while the polarization of AQP4 on the astrocyte endfeet seems to be closely related to the proliferation of reactive astrocytes.
In addition, sleep disorder is one of the common complaints in patients with head trauma. Somnolence, insomnia, and other symptoms appear in a large number of patients with TBI (Clinchot et al., 1998; Ouellet et al., 2006; Verma et al., 2007; Collen et al., 2012), suggesting that TBI damages the function of the glymphatic system by affecting patients’ normal sleep and subsequently resulting in a large accumulation of related biomarkers in the brain.
Stroke
Stroke is a group of diseases that cause brain damage due to the sudden rupture of cerebral blood vessels or vascular obstruction, including ischemic stroke and hemorrhagic stroke. In mice, both ischemic stroke and hemorrhagic stroke have been shown to lead to the blockage of CSF circulation and damage to the glymphatic pathway (Iliff et al., 2014). Goulay et al. (2017) reported a similar phenomenon in non-human primates: after subarachnoid hemorrhage (SAH), the deposition of fibrin and fibrinogen in the PVS leads to glymphatic system dysfunction and finally the deterioration of cerebral ischemia and brain edema; the application of tissue-type plasminogen activator to remove the deposits in the PVS alleviates the pathological symptoms of SAH. Liu et al. (2021) used routine MRI, diffusion tensor images scan of the brain and other methods to measure the ALPS-index and other indexes of patients with carotid plaque, which confirmed that there may be impairment of glymphatic system in patients with carotid plaque.
Post-SAH glymphatic system dysfunction significantly reduces the amount of CSF flowing into and out of the brain (Liu E. et al., 2020) and causes pathological manifestations such as vasculitis and neuroinflammation (Luo et al., 2016). The main mechanism may be the destruction of the blood brain barrier after stroke (Yang and Rosenberg, 2011). Aβ infiltrates into the CSF and brain parenchyma (Scheuner et al., 1996) and accumulates transiently in the brain parenchyma and blood vessel walls (Garcia-Alloza et al., 2011; Okamoto et al., 2012), leading to inflammation. A study has shown significantly increased AQP4 expression in the artery after SAH, but no significant change in AQP4 expression was detected in the vein, which may indicate a significant increase in the inflow of CSF from the periarterial space, while the volume of ISF expands due to the constant expression of AQP4 in the vein to eventually lead to the deterioration of brain edema (Liu E. et al., 2020). Researchers have observed increased AQP4 expression during the response to hypoxia: acute hypoxia induces AQP4 localization in cortical astrocytes and increases water permeability (Kitchen et al., 2020). This result confirms the findings reported by Mestre et al. (2020) that the entry of CSF into the brain through the glymphatic pathway is the main mechanism of edema formation and ion disturbance in the early stage of ischemic stroke. Other researchers have found that the function of the glymphatic system changes over time after ischemic stroke, and CSF inflow increases a few minutes after stroke but remains relatively slow in the following period (until 7 days) (Lin et al., 2020). Moreover, local destruction of the glymphatic system was detected around the infarct area, while the extracellular fluid in the infarct area was still toxic to neurons 7 weeks after stroke, causing long-term damage because it was not cleared in a timely manner (Zbesko et al., 2018).
Cerebral apoplexy-induced damage to the function of the glymphatic system may also lead to accelerated deposition of Aβ, tau and other proteins, which in turn trigger neurodegenerative diseases such as AD, dementia, and cognitive impairment.
Idiopathic Normal Pressure Hydrocephalus
Idiopathic normal pressure hydrocephalus (iNPH) is a neurological disease characterized by Hakeem’s triad (dementia, gait disorder, and urinary incontinence) (Lu et al., 2020). Its typical imaging findings are ventricular enlargement, wide lateral fissure, and subarachnoid stenosis (Capone et al., 2020), but intracranial pressure usually remains normal (Hakim and Adams, 1965). INPH is considered one of the most common causes of dementia and one of the risk factors for the subsequent development of AD (Ringstad et al., 2017). Different studies have used enhanced MRI to observe delayed clearance of the intrathecal CSF tracer gadobutrol in patients with iNPH (Ringstad et al., 2017; Eide and Ringstad, 2019). Yokota et al. (2019) performed diffusion tensor imaging (DTI) along the perivascular space (ALPS) and observed lower diffusivity ALPS in patients with iNPH than in the control group. Other groups reported delayed enrichment and clearance of tracers in the visual pathway of patients with iNPH (Jacobsen et al., 2020).
Glymphatic system dysfunction is a component of the pathological process of iNPH. Many possible mechanisms of glymphatic system dysfunction in iNPH pathology have been proposed. One is that excess CSF will be trapped in the dilated PVS and oppress the penetrating artery in the parenchyma of the brain to reduce its pulsation (Gallina et al., 2020). As a possible driving force of the glymphatic system, the inhibition of arterial pulsation may reduce the function of the glymphatic system. Second, decreased AQP4 expression in astrocytes of patients with iNPH mainly results in a decrease in the density of the endfeet around the blood vessels (depolarization) (Eide and Hansson, 2018; Reeves et al., 2020). As a disease related to aging, the prevalence rate of iNPH increases exponentially with age (Martín-Láez et al., 2016). The progressive depolarization of AQP4 caused by aging will further inhibit the operation of the glymphatic system (Hasan-Olive et al., 2019). Finally, patients with iNPH are often diagnosed with sleep disorders (Román et al., 2019). Obstructive sleep apnea (OSA) is a common sleep disorder associated with AD (Ooms and Ju, 2016), affecting 90.3% of patients with iNPH in one study (Román et al., 2019). Abnormal respiratory movement (intermittent airway obstruction) in people with OSA may increase intracranial venous pressure by increasing intrathoracic negative pressure and reducing venous reflux, perivenous CSF outflow and CSF-ISF exchange (Román et al., 2019), and may damage the glymphatic system while promoting the development of iNPH.
Sleep and Mental Disorders
According to recent studies, mood disorders are also associated with the glymphatic system (Xia et al., 2017b; Liu X. et al., 2020). Mood disorders are a group of mental disorders characterized by emotional imbalance, including major depressive disorder (MDD) and bipolar disorder (BD). In addition to emotional abnormalities, sleep abnormalities are also strongly associated with symptoms associated with mood disorders (Jackson et al., 2003; Lyall et al., 2018). Sleep disorder is a common complaint of patients with mood disorders, while paradoxical sleep deprivation (PSD) causes manic-like behavior in mice (Arent et al., 2015), induces mania in healthy subjects, and exacerbates manic episodes or lead to the transition from depression to mania in patients with bipolar disorder (Kaplan and Harvey, 2013). In addition, some antidepressant drugs, such as ketamine, exert an effect on sleep patterns (Duncan et al., 2017). The aforementioned evidence proves that sleep disorders play an important role in the occurrence and development of mood disorders, and because of the close relationship between sleep and the glymphatic system, we speculate that glymphatic system dysfunction is one of the possible pathological mechanisms of mood disorders. Due to altered sleep patterns or quality, glymphatic system function is abnormal, which leads to or aggravates mood disorders.
Sleep Disorder
The relationship between sleep and the glymphatic system is inseparable. During non-rapid eye movement (NREM) sleep, a consistent pattern of slow wave activity and CSF inflow is observed, supporting the exciting possibility that sleep regulates glymphatic function (Fultz et al., 2019). Siow et al. (2021) used diffusion tensor imaging-analysis along the perivascular space (DTI-ALPS) index as the MRI marker of glymphatic function, identified significant associations between glymphatic function and sleep in 84 elderly participants. However, the specific interaction between the glymphatic system and sleep remains unclear. Circadian rhythm is a natural internal process that regulates the sleep/wake cycle and repeats roughly every 24 h. Sleep/wake cycle refers to a physiological state that occurs alternately between sleep and awakening. Although previous experiments have shown that the glymphatic system clearance function is related to animal sleep/awakening states (Xie et al., 2013), recent experiments have shown that the glymphatic system is regulated by circadian rhythm rather than the sleep/wake cycle (Cai et al., 2020; Hablitz et al., 2020), which means that glymphatic system may also be affected by endogenous hormones, not just by sleep/awakening state. The redistribution of intraventricular contrast agent in conscious rats was the lowest in the light stage and the highest in the dark phase in both normal and reverse light-dark cycles, which proved the possible effect of alternating day and night cycles on the clearance function of the glymphatic system (Cai et al., 2020). The clearance function of the glymphatic system peaks at the most likely time mice sleep (noon) during the day, and the diurnal difference in glymphatic system function persists under constant light conditions, suggesting that this diurnal difference arises from endogenous regulation (Hablitz et al., 2020). At the same time, the distribution of polarized AQP4 on the endfeet of astrocytes is also related to different periods of the same day, and deletion of the AQP4 gene effectively eliminates the diurnal regulation of the CSF distribution (Hablitz et al., 2020).
Regardless of whether the glymphatic system is regulated by circadian rhythm, the sleep/wake cycle or both, circadian rhythms and sleep/wake cycles in patients with mood disorders have both been shown to be affected to varying degrees (Kalmbach et al., 2015; Bradley et al., 2017; Park et al., 2018). Symptoms of depression show diurnal patterns that are worse in the morning or at night (Rusting and Larsen, 1998), while genomic studies have further discovered the relationship between circadian rhythms and mood disorders. The circadian rhythm gene CRY2 has been proven to be related to the rapid cycling of BD (Sjöholm et al., 2010). The abnormal expression of circadian rhythm genes such as BMAL1, PER1-3, REV-ERBA, DBP, and BHLHE40/41 was also observed in the postmortem brain tissues of patients with MDD (Li et al., 2013). Sufficient studies on the sleep/wake cycle have proven the relationship between mood disorders and this cycle. As shown in our previous study, fluoxetine, a specific serotonin reuptake inhibitor widely used in the clinic, increases the activity of signal transducer and activator of transcription-3 (STAT3) in astrocytes, reverses the changes in the expression and function of nucleotide-binding domain and leucine-rich repeat protein-3 (NLRP3) inflammasomes stimulated by sleep deprivation, and then inhibits SD-induced neuroinflammation and neuronal apoptosis (Xia et al., 2017a). Sleep disorders in patients with mood disorders are likely to affect the function of the glymphatic system, but the significance of this alteration in the occurrence and development of mood disorders remains unclear.
Mood Disorders
We observed that AQP4 expression was downregulated in the cortex and hippocampus of a mouse model of chronic unpredictable mild stress (CUMS), the widespread loss distribution of AQP4 on astrocytes suggesting a disorder in the glymphatic system, and an increase in Aβ deposition was detected (Xia et al., 2017b). In addition, this disorder was improved by fluoxetine and polyunsaturated fatty acids (PUFAs) (Xia et al., 2017b; Liu X. et al., 2020). Our previous experimental results have shown that iron dextran damages the glymphatic system in the frontal cortex by increasing reactive astrogliosis, which was indicated by an increase in glial fibrillary acidic protein expression, inducing neuronal apoptosis and therefore aggravates depression-like behavior caused by CUMS (Liang et al., 2020). Decreases in the number of astrocytes and AQP4 expression were observed in other animal models of depression, such as chronic mild stress (CMS) (Gong et al., 2012) and chronic psychosocial stress (Czéh et al., 2006). Reactive astrocyte proliferation and changes in AQP4 expression are common in postmortem tissues from patients with depression, but the related experiments have produced contradictory results. Glial fibrillary acidic protein (GFAP) is one of the specific biomarkers of astrocytes. Several experimental results show that depression reduces its expression in different regions of the brain (Altshuler et al., 2010; Bernard et al., 2011; Gittins and Harrison, 2011; Cobb et al., 2016; Torres-Platas et al., 2016), but some experiments show that its expression increases (Barley et al., 2009) or it not significantly changed (Williams et al., 2013; Table 1). The expression of AQP4 in the brains of patients with MDD was shown to be downregulated by different groups [locus coeruleus (Bernard et al., 2011), hippocampus (Medina et al., 2016)]. However, in an oligonucleotide microarray analysis and qPCR study of postmortem brain tissues from patients with mood disorder, the expression of the AQP4 gene U34646 was upregulated in the prefrontal cortex (Iwamoto et al., 2004).
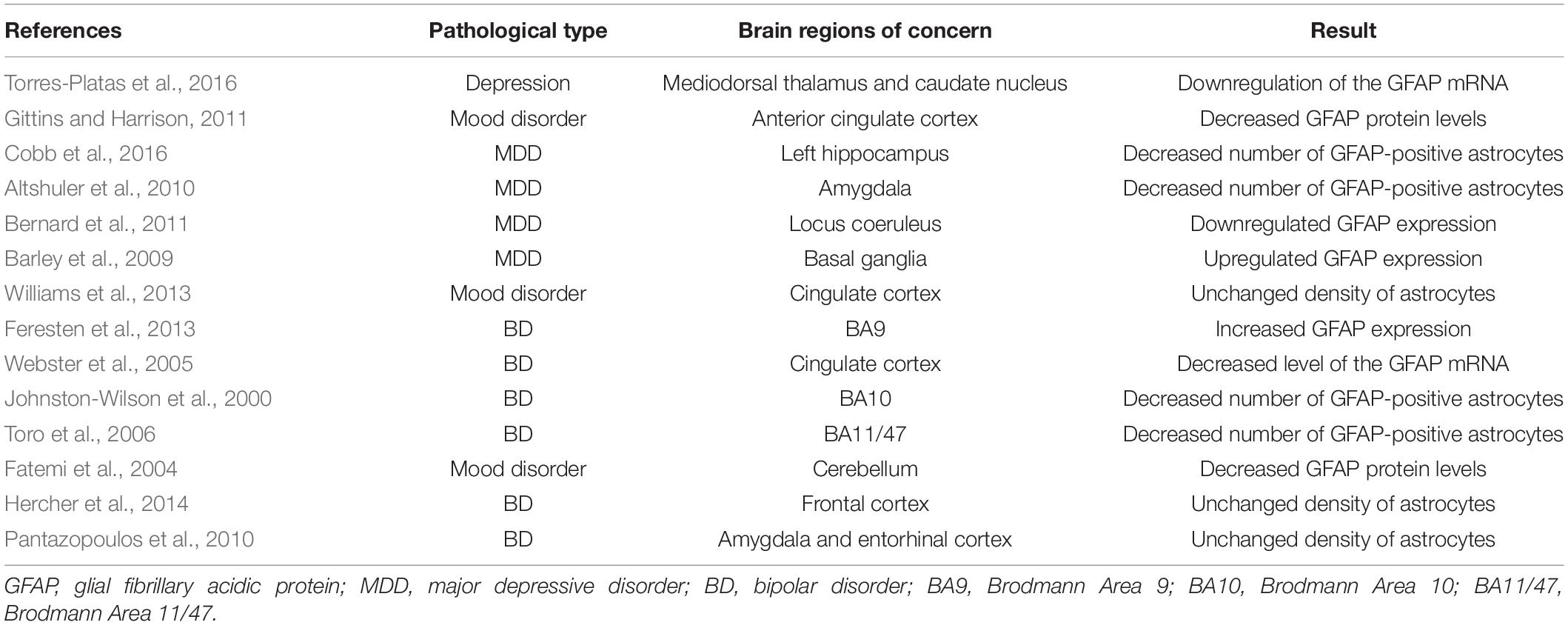
Table 1. Studies of astrocytes in postmortem tissues.
Similar to depression, astrocyte dysfunction is a very common characteristic of the occurrence and development of BD, but several experimental results show different trends. The density of astrocytes in the frontal cortex (Hercher et al., 2014), cingulate cortex (Williams et al., 2013), amygdala (Altshuler et al., 2010; Pantazopoulos et al., 2010), and entorhinal cortex (Pantazopoulos et al., 2010) did not change after death. However, some researchers have reached different conclusions: GFAP-positive astrocytes showed a decreasing trend in the BA10 (Johnston-Wilson et al., 2000) and BA11/47 (Toro et al., 2006) regions of the brain; and GFAP mRNA (Webster et al., 2005) or protein (Fatemi et al., 2004) expression decreased (see Table 1). Central nervous system-specific protein (S100β) is a calcium-binding protein that is mainly produced and secreted by astrocytes in the central nervous system. S100β is a commonly used astrocyte marker, and its level is increased in BA40 but decreased in BA9 (Dean et al., 2006). In a meta-analysis of serum S100β levels, serum levels of S100β were increased in patients with BD during the manic state (da Rosa et al., 2016).
Based on these experiments, abnormal astrocytes and AQP4 expression are very common characteristics of the pathology of mood disorders, which may be the main cause of glymphatic system dysfunction, but the contradictory experimental results require further integration and analysis.
Headache
Headache, especially migraine, is closely related to mood disorders. The prevalence rate of migraine in clinical samples of patients with BD is 34.8% (Fornaro and Stubbs, 2015). At the same time, a high coprevalence of migraine and depression has been reported (Antonaci et al., 2011). Sleep interruption is a frequently reported cause of migraine (Wang et al., 2013), and sleep is often reported as an effective method to ameliorate migraine (Singh and Sahota, 2013). Calcitonin gene-related peptide (CGRP) is the core of the pathogenesis of migraine (Edvinsson, 2017). CGRP is released not only by the afferent nerve of the trigeminal nerve innervating the meninges, pia mater and internal cerebral arteries but also by the central projection of the spinal nucleus of the trigeminal nerve at the medulla oblongata level (Messlinger et al., 1993; Lennerz et al., 2008; Eftekhari et al., 2010). Although the exact mechanism of migraine caused by CGRP remains unclear, the fact that antibodies against CGRP or its receptor have been approved for clinical treatment (Yuan et al., 2017) indicate that CGRP plays an important role in the occurrence and development of migraine. CGRP released from the trigeminal afferent cannot does not the blood brain barrier, and it may be transferred to or cleared by the glymphatic system. After dural afferents are activated by depolarized KCl, the concentration of CGRP in CSF is 5 times higher than that in plasma (Dux et al., 2017). Since the clearance of metabolites, peptides and proteins from the brain by the glymphatic system mainly occurs during sleep, we speculate that sleep disorders cause glymphatic system disorders, resulting in the accumulation of CGRP in the brain and subsequently the symptoms of migraine.
Cortical spreading inhibition (CSD) is considered the neurophysiological basis of migraine aura (Bolay et al., 2002). Schain et al. (2017) initiated CSD by either pinprick or KCl crystals (prefrontal cortex), which led to rapid and almost complete closure of the space around the artery that gradually recovered within 30 min. In this mouse model, CSD caused temporary glymphatic flow disorders, and the direct relationship between the glymphatic system and migraine was proven. This finding may supplement the pathophysiological mechanism of migraine symptoms caused by glymphatic system dysfunction.
Imaging Study
One of the main reasons for the doubt or contradictions in related research is the lack of mature imaging technology for glymphatic system. In recent years, a large number of studies have tried to use visualization technology to observe and describe the glymphatic system. The initial study used two-photon imaging observations, including fluorescent tracers and laser scanning microscopes, to discover the relationship between the glymphatic system and sleep (Xie et al., 2013) and found that alcoholism may lead to glymphatic system dysfunction (Liu Q. et al., 2020). Although optical imaging technology, such as two-photon microscopy, occupies a dominant position in the classical field, it is gradually being replaced by MRI because of its strong invasiveness.
MRI methods have been proven to be useful for the evaluation of glymphatic system in many experiments (Jiang, 2019; Taoka and Naganawa, 2020a,b). At present, the most commonly used MRI contrast agent is gadolinium-based contrast agent (GBCA). Iliff et al. (2013a) performed an intrathecal injection of the paramagnetic contrast agent GBCA and used dynamic contrast-enhanced MRI to display glymphatic function in the brains of living rats. This method characterizes the dynamic and spatial distribution of the glymphatic system in the whole brain. Because MRI has the least interference with the living body, similar methods have been used to study the glymphatic system in human subjects. A glymphatic system in humans similar to that found in the brains of rodents has been identified (Eide et al., 2018; Ringstad et al., 2018; Watts et al., 2019). Ringstad et al. (2017) used intrathecal GBCA as a CSF/ISF tracer for MRI and concluded that the glymphatic clearance rate was decreased in patients with iNPH. van de Haar et al. (2016) also used GBCA injection to confirm that the global blood-brain barrier leakage in patients with early AD was related to the decline of cognitive ability, reflecting the role of glymphatic system dysfunction in the pathological development of AD. Taoka et al. (2018) conducted animal experiments with intravenous injection of GBCA and found that the distribution of GBCA in brain tissue was affected by circadian rhythm and anesthesia. Currently, a variety of MRI methods have been developed to evaluate CSF movement, such as MRI tracer studies using the stable isotope 17O (Igarashi et al., 2014), multiple echo time (multi-TE) arterial spin labeling (ASL) MRI techniques (Ohene et al., 2019), and DTI-Alps methods to evaluate the movement of water molecules in the space around blood vessels by measuring the diffusion coefficient (Taoka et al., 2017). Abnormal glymphatic system function has been observed in the pathological processes of idiopathic normal pressure hydrocephalus (Ringstad et al., 2017; Eide and Ringstad, 2019), stroke (Gaberel et al., 2014), type 2 diabetes (Jiang et al., 2017; Yang et al., 2020), and juvenile myoclonic epilepsy (Lee et al., 2021). Different research groups have used MRI data for mathematical modeling to better understand the dynamics of glymphatic system flow (Kaur et al., 2020).
However, to date, no imaging technique has been established to directly and clearly depict the glymphatic system and neurofluid dynamics (especially in human experiments). Intrathecal injection of large doses of GBCA may cause severe gadolinium encephalopathy, potentially resulting in nausea, dyspnea, spasmodic pain of the lower extremities, and even death (Samardzic and Thamburaj, 2015; Reeves et al., 2017; Provenzano et al., 2019). Future neuroscience research is needed to develop increasingly precise measurement techniques for glymphatic system with less of an effect on normal physiological homeostasis and to improve relevant theories by obtaining quantitative data on various components of the glymphatic system as a method to obtain a deeper understanding of the relationship between the glymphatic system and various diseases.
Conclusion and Prospects for the Future
Although glymphatic system dysfunction have been observed in individuals with a variety of pathological conditions, including central nervous system diseases and mood disorders, researchers have not clearly determined whether glymphatic system damage is the cause or result of these diseases (or both) (Mestre et al., 2017). The relationship between glymphatic dysfunction and diseases, as well as the specific underlying pathophysiological processes, are not understood. In recent years, the concept of “CNS interstitial fluidopathy” has been put forward by researchers, which refers to a variety of diseases characterized by glymphatic system dysfunction or other mechanisms related to interstitial fluid dynamics (Taoka and Naganawa, 2021). The in-depth study of the relationship between these diseases and glymphatic system will not only deepen our understanding of the glymphatic system and the pathogenic mechanism of related diseases, but also promote the development of clinical treatment and prevention strategies.
In addition, some doubts persist about the glymphatic system hypothesis. First, the model based on the Navier–Stokes equation and convection-diffusion equation related to fluid motion shows that the flow resistance of water molecules through the terminal AQP4 channel on astrocytes is too large to cause a large amount of fluid flow in the interstitial space, and thus researchers have questioned the regulatory role of AQP4 in CSF flow (Jin et al., 2016). Second, regarding the driving force of the glymphatic system, Asgari et al. (2016) used mathematical modeling to prove that the arterial pulse in the tissue is too small to be the driving force for large amounts of fluid flow, while the effects of other influencing factors, such as respiration and body position, must be further clarified. Third, other processes to remove waste in the brain might exist, such as transport across the blood brain barrier (Abbott et al., 2010), degradation mediated by enzymes expressed by astrocytes (Yin et al., 2006), and macrophage uptake (Hawkes and McLaurin, 2009), but the relationship between the glymphatic system and these pathways is still unknown. Therefore, future research should focus on elucidating the driving force of the glymphatic system and whether other possible driving factors exist, as well as their effects on the glymphatic system.
Sleep and endocrine factors play important roles in regulating the brain and peripheral organs. Sleep deprivation will increase the peripheral distribution of inflammatory markers in humans and rodents (Mullington et al., 2010; Hurtado-Alvarado et al., 2013; Irwin et al., 2016), while pathological signals from the brain promote systemic inflammation through the VEGF-C/VEGFR3 signaling pathway (Esposito et al., 2019). The characteristics of brain aging include disorders of mitochondrial energy metabolism, activation of inflammatory bodies, oxidative damage, autophagy defects, inflammation, and impaired waste disposal mechanisms (Franceschi et al., 2018; Mészáros et al., 2020). The normal physiological process of aging might also cause glymphatic system dysfunction through sleep disorders and endocrine abnormalities. The change in the normal sleep pattern is directly related to aging (Mander et al., 2017), and this change may inhibit the clearance function of the glymphatic system by reducing NREM sleep (Hablitz et al., 2019) and increasing AQP4 mislocalization (Kress et al., 2014). The normal aging process also leads to chronic changes in the phenotype of microglia similar to the proinflammatory response (Tay et al., 2017), and activated microglia are part of the process inducing the formation of A1-like reactive astrocytes (Clarke et al., 2018), coupled with the cellular senescence characteristics of astrocytes themselves, such as increased levels of GFAP (Cotrina and Nedergaard, 2002; Finch, 2003) and vimentin filaments (Clarke et al., 2018); increased expression of cytokines such as tumor necrosis factor α (TNF- α), IL-1b and IL-6 (Campuzano et al., 2009); and increased accumulation of proteotoxic aggregates (Clarke et al., 2018). Lipids may also be secreted by astrocytes in an autocrine fashion, into PVSs, potentially to signal to other astrocytes, ultimately effecting ion fluxes and downstream phosphorylation pathways that themselves contribute to modulating the release or uptake of many other signaling agents (Rangroo Thrane et al., 2013). Therefore, abnormal astrocytes may affect the neural networks macroscopically. These changes may affect the normal operation of the macroscopic waste removal system in the brain through the endocrine pathway. Since sleep disorders and endocrine disorders caused by aging involve both brain and peripheral regulation and glymphatic system function, we infer that the glymphatic system plays a certain role in regulating the brain and peripheral organs. However, the role of the glymphatic system requires further study in the future.
Author Contributions
BL made the conception and design of the work, and revised the manuscript draft. DZ and XL reviewed the literature and wrote the manuscript draft. All authors contributed to the article and approved the submitted version.
Funding
This work was supported by the National Natural Science Foundation of China, BL (Grant No. 81871852), Shenyang Science and Technology Innovation Talents Project, BL (Grant No. RC210251), LiaoNing Revitalization Talents Program, BL (Grant No. XLYC1807137), the Scientific Research Foundation for Returned Scholars of Education Ministry of China, BL (Grant No. 20151098), LiaoNing Thousands Talents Program, BL (Grant No. 202078), and “ChunHui” Program of Education Ministry, BL (Grant No. 2020703).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Abbott, N. J., Patabendige, A. A., Dolman, D. E., Yusof, S. R., and Begley, D. J. (2010). Structure and function of the blood-brain barrier. Neurobiol. Dis. 37, 13–25. doi: 10.1016/j.nbd.2009.07.030
Ahmed, R. M., Newcombe, R. E., Piper, A. J., Lewis, S. J., Yee, B. J., Kiernan, M. C., et al. (2016). Sleep disorders and respiratory function in amyotrophic lateral sclerosis. Sleep Med. Rev. 26, 33–42. doi: 10.1016/j.smrv.2015.05.007
Albrecht, U. (2013). Circadian clocks and mood-related behaviors. Handb. Exp. Pharmacol. 217, 227–239. doi: 10.1007/978-3-642-25950-0_9
Altshuler, L. L., Abulseoud, O. A., Foland-Ross, L., Bartzokis, G., Chang, S., Mintz, J., et al. (2010). Amygdala astrocyte reduction in subjects with major depressive disorder but not bipolar disorder. Bipolar Disord. 12, 541–549. doi: 10.1111/j.1399-5618.2010.00838.x
Antonaci, F., Nappi, G., Galli, F., Manzoni, G. C., Calabresi, P., and Costa, A. (2011). Migraine and psychiatric comorbidity: a review of clinical findings. J. Headache Pain 12, 115–125. doi: 10.1007/s10194-010-0282-4
Arent, C. O., Valvassori, S. S., Steckert, A. V., Resende, W. R., Dal-Pont, G. C., Lopes-Borges, J., et al. (2015). The effects of n-acetylcysteine and/or deferoxamine on manic-like behavior and brain oxidative damage in mice submitted to the paradoxal sleep deprivation model of mania. J. Psychiatr. Res. 65, 71–79. doi: 10.1016/j.jpsychires.2015.04.011
Armstrong, R. A. (2009). The molecular biology of senile plaques and neurofibrillary tangles in Alzheimer’s disease. Folia Neuropathol. 47, 289–299.
Asgari, M., de Zélicourt, D., and Kurtcuoglu, V. (2016). Glymphatic solute transport does not require bulk flow. Sci. Rep. 6:38635. doi: 10.1038/srep38635
Barber, A., and Dashtipour, K. (2012). Sleep disturbances in Parkinson’s disease with emphasis on rapid eye movement sleep behavior disorder. Int. J. Neurosci. 122, 407–412. doi: 10.3109/00207454.2012.677882
Barley, K., Dracheva, S., and Byne, W. (2009). Subcortical oligodendrocyte- and astrocyte-associated gene expression in subjects with schizophrenia, major depression and bipolar disorder. Schizophr. Res. 112, 54–64. doi: 10.1016/j.schres.2009.04.019
Bernard, R., Kerman, I. A., Thompson, R. C., Jones, E. G., Bunney, W. E., Barchas, J. D., et al. (2011). Altered expression of glutamate signaling, growth factor, and glia genes in the locus coeruleus of patients with major depression. Mol. Psychiatry 16, 634–646. doi: 10.1038/mp.2010.44
Berridge, C. W., and Waterhouse, B. D. (2003). The locus coeruleus-noradrenergic system: modulation of behavioral state and state-dependent cognitive processes. Brain Res. Brain Res. Rev. 42, 33–84. doi: 10.1016/s0165-0173(03)00143-7
Bertel, O., Malessa, S., Sluga, E., and Hornykiewicz, O. (1991). Amyotrophic lateral sclerosis: changes of noradrenergic and serotonergic transmitter systems in the spinal cord. Brain Res. 566, 54–60. doi: 10.1016/0006-8993(91)91680-y
Binder, L. I., Guillozet-Bongaarts, A. L., Garcia-Sierra, F., and Berry, R. W. (2005). Tau, tangles, and Alzheimer’s disease. Biochim. Biophys. Acta 1739, 216–223. doi: 10.1016/j.bbadis.2004.08.014
Bobela, W., Aebischer, P., and Schneider, B. L. (2015). A lpha-synuclein as a mediator in the interplay between aging and Parkinson’s disease. Biomolecules 5, 2675–2700. doi: 10.3390/biom5042675
Bohnen, N. I., and Hu, M. T. M. (2019). Sleep disturbance as potential risk and progression factor for Parkinson’s disease. J. Parkinsons Dis. 9, 603–614. doi: 10.3233/jpd-191627
Bolay, H., Reuter, U., Dunn, A. K., Huang, Z., Boas, D. A., and Moskowitz, M. A. (2002). Intrinsic brain activity triggers trigeminal meningeal afferents in a migraine model. Nat. Med. 8, 136–142. doi: 10.1038/nm0202-136
Bolte, A. C., and Lukens, J. R. (2021). Neuroimmune cleanup crews in brain injury. Trends Immunol. 42, 480–494. doi: 10.1016/j.it.2021.04.003
Bradley, A. J., Webb-Mitchell, R., Hazu, A., Slater, N., Middleton, B., Gallagher, P., et al. (2017). Sleep and circadian rhythm disturbance in bipolar disorder. Psychol. Med. 47, 1678–1689. doi: 10.1017/s0033291717000186
Cai, X., Qiao, J., Kulkarni, P., Harding, I. C., Ebong, E., and Ferris, C. F. (2020). Imaging the effect of the circadian light-dark cycle on the glymphatic system in awake rats. Proc. Natl. Acad. Sci. U.S.A. 117, 668–676. doi: 10.1073/pnas.1914017117
Campuzano, O., Castillo-Ruiz, M. M., Acarin, L., Castellano, B., and Gonzalez, B. (2009). Increased levels of proinflammatory cytokines in the aged rat brain attenuate injury-induced cytokine response after excitotoxic damage. J. Neurosci. Res. 87, 2484–2497. doi: 10.1002/jnr.22074
Capone, P. M., Bertelson, J. A., and Ajtai, B. (2020). Neuroimaging of normal pressure hydrocephalus and hydrocephalus. Neurol. Clin. 38, 171–183. doi: 10.1016/j.ncl.2019.09.003
Chen, G. F., Xu, T. H., Yan, Y., Zhou, Y. R., Jiang, Y., Melcher, K., et al. (2017). Amyloid beta: structure, biology and structure-based therapeutic development. Acta Pharmacol. Sin. 38, 1205–1235. doi: 10.1038/aps.2017.28
Chiang, P. L., Chen, H. L., Lu, C. H., Chen, Y. S., Chou, K. H., Hsu, T. W., et al. (2018). Interaction of systemic oxidative stress and mesial temporal network degeneration in Parkinson’s disease with and without cognitive impairment. J. Neuroinflammation 15:281. doi: 10.1186/s12974-018-1317-z
Christensen, J., Wright, D. K., Yamakawa, G. R., Shultz, S. R., and Mychasiuk, R. (2020). Repetitive mild traumatic brain injury alters glymphatic clearance rates in limbic structures of adolescent female rats. Sci. Rep. 10:6254. doi: 10.1038/s41598-020-63022-7
Clarke, L. E., Liddelow, S. A., Chakraborty, C., Münch, A. E., Heiman, M., and Barres, B. A. (2018). Normal aging induces A1-like astrocyte reactivity. Proc. Natl. Acad. Sci. U.S.A. 115, E1896–E1905. doi: 10.1073/pnas.1800165115
Clinchot, D. M., Bogner, J., Mysiw, W. J., Fugate, L., and Corrigan, J. (1998). Defining sleep disturbance after brain injury. Am. J. Phys. Med. Rehabil. 77, 291–295. doi: 10.1097/00002060-199807000-00006
Cobb, J. A., O’Neill, K., Milner, J., Mahajan, G. J., Lawrence, T. J., May, W. L., et al. (2016). Density of GFAP-immunoreactive astrocytes is decreased in left hippocampi in major depressive disorder. Neuroscience 316, 209–220. doi: 10.1016/j.neuroscience.2015.12.044
Collen, J., Orr, N., Lettieri, C. J., Carter, K., and Holley, A. B. (2012). Sleep disturbances among soldiers with combat-related traumatic brain injury. Chest 142, 622–630. doi: 10.1378/chest.11-1603
Cotrina, M. L., and Nedergaard, M. (2002). Astrocytes in the aging brain. J. Neurosci. Res. 67, 1–10. doi: 10.1002/jnr.10121
Czéh, B., Simon, M., Schmelting, B., Hiemke, C., and Fuchs, E. (2006). Astroglial plasticity in the hippocampus is affected by chronic psychosocial stress and concomitant fluoxetine treatment. Neuropsychopharmacology 31, 1616–1626. doi: 10.1038/sj.npp.1300982
da Rosa, M. I., Simon, C., Grande, A. J., Barichello, T., Oses, J. P., and Quevedo, J. (2016). Serum S100B in manic bipolar disorder patients: systematic review and meta-analysis. J. Affect. Disord. 206, 210–215. doi: 10.1016/j.jad.2016.07.030
Dai, J., Lin, W., Zheng, M., Liu, Q., He, B., Luo, C., et al. (2017). Alterations in AQP4 expression and polarization in the course of motor neuron degeneration in SOD1G93A mice. Mol. Med. Rep. 16, 1739–1746. doi: 10.3892/mmr.2017.6786
Dean, B., Gray, L., and Scarr, E. (2006). Regionally specific changes in levels of cortical S100beta in bipolar 1 disorder but not schizophrenia. Aust. N. Z. J. Psychiatry 40, 217–224. doi: 10.1080/j.1440-1614.2006.01777.x
De-Paula, V. J., Radanovic, M., Diniz, B. S., and Forlenza, O. V. (2012). Alzheimer’s disease. Subcell Biochem. 65, 329–352. doi: 10.1007/978-94-007-5416-4_14
Dickson, D. W., Fujishiro, H., Orr, C., DelleDonne, A., Josephs, K. A., Frigerio, R., et al. (2009). Neuropathology of non-motor features of Parkinson disease. Parkinsonism Relat. Disord. 15(Suppl. 3), S1–S5. doi: 10.1016/s1353-8020(09)70769-2
Duncan, W. C. Jr., Slonena, E., Hejazi, N. S., Brutsche, N., Yu, K. C., Park, L., et al. (2017). Motor-activity markers of circadian timekeeping are related to Ketamine’s rapid antidepressant properties. Biol. Psychiatry 82, 361–369. doi: 10.1016/j.biopsych.2017.03.011
Dux, M., Will, C., Eberhardt, M., Fischer, M. J. M., and Messlinger, K. (2017). Stimulation of rat cranial dura mater with potassium chloride causes CGRP release into the cerebrospinal fluid and increases medullary blood flow. Neuropeptides 64, 61–68. doi: 10.1016/j.npep.2017.02.080
Edvinsson, L. (2017). The trigeminovascular pathway: role of CGRP and CGRP receptors in migraine. Headache 57(Suppl. 2), 47–55. doi: 10.1111/head.13081
Eftekhari, S., Salvatore, C. A., Calamari, A., Kane, S. A., Tajti, J., and Edvinsson, L. (2010). Differential distribution of calcitonin gene-related peptide and its receptor components in the human trigeminal ganglion. Neuroscience 169, 683–696. doi: 10.1016/j.neuroscience.2010.05.016
Eide, P. K., and Hansson, H. A. (2018). Astrogliosis and impaired aquaporin-4 and dystrophin systems in idiopathic normal pressure hydrocephalus. Neuropathol. Appl. Neurobiol. 44, 474–490. doi: 10.1111/nan.12420
Eide, P. K., and Ringstad, G. (2019). Delayed clearance of cerebrospinal fluid tracer from entorhinal cortex in idiopathic normal pressure hydrocephalus: a glymphatic magnetic resonance imaging study. J. Cereb. Blood Flow Metab. 39, 1355–1368. doi: 10.1177/0271678x18760974
Eide, P. K., Vatnehol, S. A. S., Emblem, K. E., and Ringstad, G. (2018). Magnetic resonance imaging provides evidence of glymphatic drainage from human brain to cervical lymph nodes. Sci. Rep. 8:7194. doi: 10.1038/s41598-018-25666-4
Esposito, E., Ahn, B. J., Shi, J., Nakamura, Y., Park, J. H., Mandeville, E. T., et al. (2019). Brain-to-cervical lymph node signaling after stroke. Nat. Commun. 10:5306. doi: 10.1038/s41467-019-13324-w
Fatemi, S. H., Laurence, J. A., Araghi-Niknam, M., Stary, J. M., Schulz, S. C., Lee, S., et al. (2004). Glial fibrillary acidic protein is reduced in cerebellum of subjects with major depression, but not schizophrenia. Schizophr. Res. 69, 317–323. doi: 10.1016/j.schres.2003.08.014
Feresten, A. H., Barakauskas, V., Ypsilanti, A., Barr, A. M., and Beasley, C. L. (2013). Increased expression of glial fibrillary acidic protein in prefrontal cortex in psychotic illness. Schizophr. Res. 150, 252–257. doi: 10.1016/j.schres.2013.07.024
Finch, C. E. (2003). Neurons, glia, and plasticity in normal brain aging. Neurobiol. Aging 24(Suppl. 1), S123–S127. doi: 10.1016/s0197-4580(03)00051-4
Fornaro, M., and Stubbs, B. (2015). A meta-analysis investigating the prevalence and moderators of migraines among people with bipolar disorder. J. Affect. Disord. 178, 88–97. doi: 10.1016/j.jad.2015.02.032
Franceschi, C., Garagnani, P., Parini, P., Giuliani, C., and Santoro, A. (2018). Inflammaging: a new immune-metabolic viewpoint for age-related diseases. Nat. Rev. Endocrinol. 14, 576–590. doi: 10.1038/s41574-018-0059-4
Fultz, N. E., Bonmassar, G., Setsompop, K., Stickgold, R. A., Rosen, B. R., Polimeni, J. R., et al. (2019). Coupled electrophysiological, hemodynamic, and cerebrospinal fluid oscillations in human sleep. Science 366, 628–631. doi: 10.1126/science.aax5440
Gabbita, S. P., Scheff, S. W., Menard, R. M., Roberts, K., Fugaccia, I., and Zemlan, F. P. (2005). Cleaved-tau: a biomarker of neuronal damage after traumatic brain injury. J. Neurotrauma 22, 83–94. doi: 10.1089/neu.2005.22.83
Gaberel, T., Gakuba, C., Goulay, R., Martinez De Lizarrondo, S., Hanouz, J. L., Emery, E., et al. (2014). Impaired glymphatic perfusion after strokes revealed by contrast-enhanced MRI: a new target for fibrinolysis? Stroke 45, 3092–3096. doi: 10.1161/strokeaha.114.006617
Gallina, P., Porfirio, B., and Lolli, F. (2020). iNPH as a ‘2-hit’ intracranial hydrodynamic derangement disease. Trends Mol Med 26, 531–532. doi: 10.1016/j.molmed.2020.04.002
Garcia-Alloza, M., Gregory, J., Kuchibhotla, K. V., Fine, S., Wei, Y., Ayata, C., et al. (2011). Cerebrovascular lesions induce transient β-amyloid deposition. Brain 134(Pt 12), 3697–3707. doi: 10.1093/brain/awr300
Geuens, T., Bouhy, D., and Timmerman, V. (2016). The hnRNP family: insights into their role in health and disease. Hum. Genet. 135, 851–867. doi: 10.1007/s00439-016-1683-5
Gittins, R. A., and Harrison, P. J. (2011). A morphometric study of glia and neurons in the anterior cingulate cortex in mood disorder. J. Affect. Disord. 133, 328–332. doi: 10.1016/j.jad.2011.03.042
Giza, C. C., and Hovda, D. A. (2014). The new neurometabolic cascade of concussion. Neurosurgery 75(Suppl 4), S24–S33. doi: 10.1227/neu.0000000000000505
Gong, Y., Sun, X. L., Wu, F. F., Su, C. J., Ding, J. H., and Hu, G. (2012). Female early adult depression results in detrimental impacts on the behavioral performance and brain development in offspring. CNS Neurosci. Ther. 18, 461–470. doi: 10.1111/j.1755-5949.2012.00324.x
Goulay, R., Aron Badin, R., Flament, J., Emery, E., Hantraye, P., Vivien, D., et al. (2018). Cerebrospinal fluid leakage after posterior fossa surgery may impair brain metabolite clearance. Neurochirurgie 64, 422–424. doi: 10.1016/j.neuchi.2018.08.003
Goulay, R., Flament, J., Gauberti, M., Naveau, M., Pasquet, N., Gakuba, C., et al. (2017). Subarachnoid hemorrhage severely impairs brain parenchymal cerebrospinal fluid circulation in nonhuman primate. Stroke 48, 2301–2305. doi: 10.1161/strokeaha.117.017014
Grinberg, L. T., Rueb, U., Alho, A. T., and Heinsen, H. (2010). Brainstem pathology and non-motor symptoms in PD. J. Neurol. Sci. 289, 81–88. doi: 10.1016/j.jns.2009.08.021
Hablitz, L. M., Plá, V., Giannetto, M., Vinitsky, H. S., Stæger, F. F., Metcalfe, T., et al. (2020). Circadian control of brain glymphatic and lymphatic fluid flow. Nat. Commun. 11:4411. doi: 10.1038/s41467-020-18115-2
Hablitz, L. M., Vinitsky, H. S., Sun, Q., Stæger, F. F., Sigurdsson, B., Mortensen, K. N., et al. (2019). Increased glymphatic influx is correlated with high EEG delta power and low heart rate in mice under anesthesia. Sci. Adv. 5:eaav5447. doi: 10.1126/sciadv.aav5447
Hakim, S., and Adams, R. D. (1965). The special clinical problem of symptomatic hydrocephalus with normal cerebrospinal fluid pressure. Observations on cerebrospinal fluid hydrodynamics. J. Neurol. Sci. 2, 307–327. doi: 10.1016/0022-510x(65)90016-x
Harrison, I. F., Ismail, O., Machhada, A., Colgan, N., Ohene, Y., Nahavandi, P., et al. (2020). Impaired glymphatic function and clearance of tau in an Alzheimer’s disease model. Brain 143, 2576–2593. doi: 10.1093/brain/awaa179
Hasan-Olive, M. M., Enger, R., Hansson, H. A., Nagelhus, E. A., and Eide, P. K. (2019). Loss of perivascular aquaporin-4 in idiopathic normal pressure hydrocephalus. Glia 67, 91–100. doi: 10.1002/glia.23528
Hawkes, C. A., and McLaurin, J. (2009). Selective targeting of perivascular macrophages for clearance of beta-amyloid in cerebral amyloid angiopathy. Proc. Natl. Acad. Sci. U.S.A. 106, 1261–1266. doi: 10.1073/pnas.0805453106
Henderson, M. X., Trojanowski, J. Q., and Lee, V. M. (2019). α-Synuclein pathology in Parkinson’s disease and related α-synucleinopathies. Neurosci. Lett. 709:134316. doi: 10.1016/j.neulet.2019.134316
Hercher, C., Chopra, V., and Beasley, C. L. (2014). Evidence for morphological alterations in prefrontal white matter glia in schizophrenia and bipolar disorder. J. Psychiatry Neurosci. 39, 376–385. doi: 10.1503/jpn.130277
Hirose, M., Asano, M., Watanabe-Matsumoto, S., Yamanaka, K., Abe, Y., Yasui, M., et al. (2021). Stagnation of glymphatic interstitial fluid flow and delay in waste clearance in the SOD1-G93A mouse model of ALS. Neurosci. Res. 171, 74–82. doi: 10.1016/j.neures.2020.10.006
Hoshi, A., Tsunoda, A., Tada, M., Nishizawa, M., Ugawa, Y., and Kakita, A. (2017). Expression of Aquaporin 1 and Aquaporin 4 in the temporal neocortex of patients with Parkinson’s disease. Brain Pathol. 27, 160–168. doi: 10.1111/bpa.12369
Hurtado-Alvarado, G., Pavón, L., Castillo-García, S. A., Hernández, M. E., Domínguez-Salazar, E., Velázquez-Moctezuma, J., et al. (2013). Sleep loss as a factor to induce cellular and molecular inflammatory variations. Clin. Dev. Immunol. 2013:801341. doi: 10.1155/2013/801341
Igarashi, H., Tsujita, M., Kwee, I. L., and Nakada, T. (2014). Water influx into cerebrospinal fluid is primarily controlled by aquaporin-4, not by aquaporin-1: 17O JJVCPE MRI study in knockout mice. Neuroreport 25, 39–43. doi: 10.1097/wnr.0000000000000042
Iliff, J. J., Chen, M. J., Plog, B. A., Zeppenfeld, D. M., Soltero, M., Yang, L., et al. (2014). Impairment of glymphatic pathway function promotes tau pathology after traumatic brain injury. J. Neurosci. 34, 16180–16193. doi: 10.1523/jneurosci.3020-14.2014
Iliff, J. J., Lee, H., Yu, M., Feng, T., Logan, J., Nedergaard, M., et al. (2013a). Brain-wide pathway for waste clearance captured by contrast-enhanced MRI. J. Clin. Invest. 123, 1299–1309. doi: 10.1172/jci67677
Iliff, J. J., Wang, M., Zeppenfeld, D. M., Venkataraman, A., Plog, B. A., Liao, Y., et al. (2013b). Cerebral arterial pulsation drives paravascular CSF-interstitial fluid exchange in the murine brain. J. Neurosci. 33, 18190–18199. doi: 10.1523/jneurosci.1592-13.2013
Iliff, J. J., Wang, M., Liao, Y., Plogg, B. A., Peng, W., Gundersen, G. A., et al. (2012). A paravascular pathway facilitates CSF flow through the brain parenchyma and the clearance of interstitial solutes, including amyloid β. Sci. Transl. Med. 4:147ra111. doi: 10.1126/scitranslmed.3003748
Imbesi, M., Yildiz, S., Dirim Arslan, A., Sharma, R., Manev, H., and Uz, T. (2009). Dopamine receptor-mediated regulation of neuronal “clock” gene expression. Neuroscience 158, 537–544. doi: 10.1016/j.neuroscience.2008.10.044
Inglese, M., Bomsztyk, E., Gonen, O., Mannon, L. J., Grossman, R. I., and Rusinek, H. (2005). Dilated perivascular spaces: hallmarks of mild traumatic brain injury. AJNR Am. J. Neuroradiol. 26, 719–724.
Irwin, M. R., Olmstead, R., and Carroll, J. E. (2016). Sleep disturbance, sleep duration, and inflammation: a systematic review and meta-analysis of cohort studies and experimental sleep deprivation. Biol. Psychiatry 80, 40–52. doi: 10.1016/j.biopsych.2015.05.014
Isaac, S. O., and Berridge, C. W. (2003). Wake-promoting actions of dopamine D1 and D2 receptor stimulation. J. Pharmacol. Exp. Ther. 307, 386–394. doi: 10.1124/jpet.103.053918
Iwamoto, K., Kakiuchi, C., Bundo, M., Ikeda, K., and Kato, T. (2004). Molecular characterization of bipolar disorder by comparing gene expression profiles of postmortem brains of major mental disorders. Mol. Psychiatry 9, 406–416. doi: 10.1038/sj.mp.4001437
Jackson, A., Cavanagh, J., and Scott, J. (2003). A systematic review of manic and depressive prodromes. J. Affect. Disord. 74, 209–217. doi: 10.1016/s0165-0327(02)00266-5
Jacobsen, H. H., Sandell, T., Jørstad, ØK., Moe, M. C., Ringstad, G., and Eide, P. K. (2020). In vivo evidence for impaired glymphatic function in the visual pathway of patients with normal pressure hydrocephalus. Invest. Ophthalmol. Vis. Sci. 61:24. doi: 10.1167/iovs.61.13.24
Jessen, N. A., Munk, A. S., Lundgaard, I., and Nedergaard, M. (2015). The glymphatic system: a beginner’s guide. Neurochem. Res. 40, 2583–2599. doi: 10.1007/s11064-015-1581-6
Jiang, Q. (2019). MRI and glymphatic system. Stroke Vasc. Neurol. 4, 75–77. doi: 10.1136/svn-2018-000197
Jiang, Q., Zhang, L., Ding, G., Davoodi-Bojd, E., Li, Q., Li, L., et al. (2017). Impairment of the glymphatic system after diabetes. J. Cereb. Blood Flow Metab. 37, 1326–1337. doi: 10.1177/0271678x16654702
Jin, B. J., Smith, A. J., and Verkman, A. S. (2016). Spatial model of convective solute transport in brain extracellular space does not support a “glymphatic” mechanism. J. Gen. Physiol. 148, 489–501. doi: 10.1085/jgp.201611684
Johnston-Wilson, N. L., Sims, C. D., Hofmann, J. P., Anderson, L., Shore, A. D., Torrey, E. F., et al. (2000). Disease-specific alterations in frontal cortex brain proteins in schizophrenia, bipolar disorder, and major depressive disorder. The stanley neuropathology consortium. Mol. Psychiatry 5, 142–149. doi: 10.1038/sj.mp.4000696
Kalaitzakis, M. E., Gentleman, S. M., and Pearce, R. K. (2013). Disturbed sleep in Parkinson’s disease: anatomical and pathological correlates. Neuropathol. Appl. Neurobiol. 39, 644–653. doi: 10.1111/nan.12024
Kalaitzakis, M. E., and Pearce, R. K. (2009). The morbid anatomy of dementia in Parkinson’s disease. Acta Neuropathol. 118, 587–598. doi: 10.1007/s00401-009-0597-x
Kalmbach, D. A., Pillai, V., Cheng, P., Arnedt, J. T., and Drake, C. L. (2015). Shift work disorder, depression, and anxiety in the transition to rotating shifts: the role of sleep reactivity. Sleep Med. 16, 1532–1538. doi: 10.1016/j.sleep.2015.09.007
Kaplan, K. A., and Harvey, A. G. (2013). Behavioral treatment of insomnia in bipolar disorder. Am. J. Psychiatry 170, 716–720. doi: 10.1176/appi.ajp.2013.12050708
Kaur, J., Davoodi-Bojd, E., Fahmy, L. M., Zhang, L., Ding, G., Hu, J., et al. (2020). Magnetic resonance imaging and modeling of the glymphatic system. Diagnostics (Basel) 10:344. doi: 10.3390/diagnostics10060344
Kiernan, M. C., Vucic, S., Cheah, B. C., Turner, M. R., Eisen, A., Hardiman, O., et al. (2011). Amyotrophic lateral sclerosis. Lancet 377, 942–955. doi: 10.1016/s0140-6736(10)61156-7
Kitchen, P., Salman, M. M., Halsey, A. M., Clarke-Bland, C., MacDonald, J. A., Ishida, H., et al. (2020). Targeting Aquaporin-4 subcellular localization to treat central nervous system edema. Cell 181, 784–799.e19. doi: 10.1016/j.cell.2020.03.037
Korogod, N., Petersen, C. C., and Knott, G. W. (2015). Ultrastructural analysis of adult mouse neocortex comparing aldehyde perfusion with cryo fixation. Elife 4:e05793. doi: 10.7554/eLife.05793
Kotajima, F., Meadows, G. E., Morrell, M. J., and Corfield, D. R. (2005). Cerebral blood flow changes associated with fluctuations in alpha and theta rhythm during sleep onset in humans. J. Physiol. 568(Pt 1), 305–313. doi: 10.1113/jphysiol.2005.092577
Kovacs, G. G. (2016). Molecular pathological classification of neurodegenerative diseases: turning towards precision medicine. Int. J. Mol. Sci. 17:189. doi: 10.3390/ijms17020189
Kovacs, G. G. (2019). Molecular pathology of neurodegenerative diseases: principles and practice. J. Clin. Pathol. 72, 725–735. doi: 10.1136/jclinpath-2019-205952
Kovacs, G. G., Milenkovic, I. J., Preusser, M., and Budka, H. (2008). Nigral burden of alpha-synuclein correlates with striatal dopamine deficit. Mov. Disord. 23, 1608–1612. doi: 10.1002/mds.22207
Kress, B. T., Iliff, J. J., Xia, M., Wang, M., Wei, H. S., Zeppenfeld, D., et al. (2014). Impairment of paravascular clearance pathways in the aging brain. Ann. Neurol. 76, 845–861. doi: 10.1002/ana.24271
Küppers, E., Gleiser, C., Brito, V., Wachter, B., Pauly, T., Hirt, B., et al. (2008). AQP4 expression in striatal primary cultures is regulated by dopamine–implications for proliferation of astrocytes. Eur. J. Neurosci. 28, 2173–2182. doi: 10.1111/j.1460-9568.2008.06531.x
Kyrtsos, C. R., and Baras, J. S. (2015). Modeling the role of the glymphatic pathway and cerebral blood vessel properties in Alzheimer’s disease pathogenesis. PLoS One 10:e0139574. doi: 10.1371/journal.pone.0139574
Lee, H. J., Lee, D. A., Shin, K. J., and Park, K. M. (2021). Glymphatic system dysfunction in patients with juvenile myoclonic epilepsy. J. Neurol. 269, 2133–2139. doi: 10.1007/s00415-021-10799-w
Lennerz, J. K., Rühle, V., Ceppa, E. P., Neuhuber, W. L., Bunnett, N. W., Grady, E. F., et al. (2008). Calcitonin receptor-like receptor (CLR), receptor activity-modifying protein 1 (RAMP1), and calcitonin gene-related peptide (CGRP) immunoreactivity in the rat trigeminovascular system: differences between peripheral and central CGRP receptor distribution. J. Comp. Neurol. 507, 1277–1299. doi: 10.1002/cne.21607
Li, J. Z., Bunney, B. G., Meng, F., Hagenauer, M. H., Walsh, D. M., Vawter, M. P., et al. (2013). Circadian patterns of gene expression in the human brain and disruption in major depressive disorder. Proc. Natl. Acad. Sci. U.S.A. 110, 9950–9955. doi: 10.1073/pnas.1305814110
Liang, S., Lu, Y., Li, Z., Li, S., Chen, B., Zhang, M., et al. (2020). Iron aggravates the depressive phenotype of stressed mice by compromising the glymphatic system. Neurosci. Bull. 36, 1542–1546. doi: 10.1007/s12264-020-00539-x
Lin, L., Hao, X., Li, C., Sun, C., Wang, X., Yin, L., et al. (2020). Impaired glymphatic system in secondary degeneration areas after ischemic stroke in rats. J. Stroke Cerebrovasc. Dis. 29:104828. doi: 10.1016/j.jstrokecerebrovasdis.2020.104828
Liu, E., Peng, X., Ma, H., Zhang, Y., Yang, X., Zhang, Y., et al. (2020). The involvement of Aquaporin-4 in the interstitial fluid drainage impairment following subarachnoid hemorrhage. Front. Aging Neurosci. 12:611494. doi: 10.3389/fnagi.2020.611494
Liu, H., Yang, S., He, W., Liu, X., Sun, S., Wang, S., et al. (2021). Associations among diffusion tensor image along the perivascular space (DTI-ALPS), enlarged perivascular space (ePVS), and cognitive functions in asymptomatic patients with carotid plaque. Front. Neurol. 12:789918. doi: 10.3389/fneur.2021.789918
Liu, Q., Yan, L., Huang, M., Zeng, H., Satyanarayanan, S. K., Shi, Z., et al. (2020). Experimental alcoholism primes structural and functional impairment of the glymphatic pathway. Brain Behav. Immun. 85, 106–119. doi: 10.1016/j.bbi.2019.06.029
Liu, X., Hao, J., Yao, E., Cao, J., Zheng, X., Yao, D., et al. (2020). Polyunsaturated fatty acid supplement alleviates depression-incident cognitive dysfunction by protecting the cerebrovascular and glymphatic systems. Brain Behav. Immun. 89, 357–370. doi: 10.1016/j.bbi.2020.07.022
Lu, V. M., Kerezoudis, P., Patel, N. P., Jones, D. T., Cutsforth-Gregory, J. K., Graff-Radford, J., et al. (2020). Our efforts in understanding normal pressure hydrocephalus: learning from the 100 most cited articles by bibliometric analysis. World Neurosurg. 137, 429–434e.13. doi: 10.1016/j.wneu.2020.02.021
Lundgaard, I., Li, B., Xie, L., Kang, H., Sanggaard, S., Haswell, J. D., et al. (2015). Direct neuronal glucose uptake heralds activity-dependent increases in cerebral metabolism. Nat. Commun. 6:6807. doi: 10.1038/ncomms7807
Luo, C., Yao, X., Li, J., He, B., Liu, Q., Ren, H., et al. (2016). Paravascular pathways contribute to vasculitis and neuroinflammation after subarachnoid hemorrhage independently of glymphatic control. Cell Death Dis. 7:e2160. doi: 10.1038/cddis.2016.63
Lyall, L. M., Wyse, C. A., Graham, N., Ferguson, A., Lyall, D. M., Cullen, B., et al. (2018). Association of disrupted circadian rhythmicity with mood disorders, subjective wellbeing, and cognitive function: a cross-sectional study of 91 105 participants from the UK Biobank. Lancet Psychiatry 5, 507–514. doi: 10.1016/s2215-0366(18)30139-1
Mader, S., and Brimberg, L. (2019). Aquaporin-4 water channel in the brain and its implication for health and disease. Cells 8:90. doi: 10.3390/cells8020090
Mander, B. A., Winer, J. R., and Walker, M. P. (2017). Sleep and human aging. Neuron 94, 19–36. doi: 10.1016/j.neuron.2017.02.004
Martín-Láez, R., Caballero-Arzapalo, H., Valle-San Román, N., López-Menéndez, L., Arango-Lasprilla, J. C., and Vázquez-Barquero, A. (2016). Incidence of idiopathic normal-pressure hydrocephalus in Northern Spain. World Neurosurg. 87, 298–310. doi: 10.1016/j.wneu.2015.10.069
Medina, A., Watson, S. J., Bunney, W. Jr., Myers, R. M., Schatzberg, A., Barchas, J., et al. (2016). Evidence for alterations of the glial syncytial function in major depressive disorder. J. Psychiatr Res. 72, 15–21. doi: 10.1016/j.jpsychires.2015.10.010
Meng, Y., Abrahao, A., Heyn, C. C., Bethune, A. J., Huang, Y., Pople, C. B., et al. (2019). Glymphatics visualization after focused ultrasound-induced blood-brain barrier opening in humans. Ann. Neurol. 86, 975–980. doi: 10.1002/ana.25604
Messlinger, K., Hanesch, U., Baumgärtel, M., Trost, B., and Schmidt, R. F. (1993). Innervation of the dura mater encephali of cat and rat: ultrastructure and calcitonin gene-related peptide-like and substance P-like immunoreactivity. Anat. Embryol. (Berl.) 188, 219–237. doi: 10.1007/bf00188214
Mestre, H., Du, T., Sweeney, A. M., Liu, G., Samson, A. J., Peng, W., et al. (2020). Cerebrospinal fluid influx drives acute ischemic tissue swelling. Science 367:eaax7171. doi: 10.1126/science.aax7171
Mestre, H., Hablitz, L. M., Xavier, A. L., Feng, W., Zou, W., Pu, T., et al. (2018). Aquaporin-4-dependent glymphatic solute transport in the rodent brain. Elife 7:e40070. doi: 10.7554/eLife.40070
Mestre, H., Kostrikov, S., Mehta, R. I., and Nedergaard, M. (2017). Perivascular spaces, glymphatic dysfunction, and small vessel disease. Clin. Sci. (Lond.) 131, 2257–2274. doi: 10.1042/cs20160381
Mészáros, Á, Molnár, K., Nógrádi, B., Hernádi, Z., Nyúl-Tóth, Á, Wilhelm, I., et al. (2020). Neurovascular inflammaging in health and disease. Cells 9:1614. doi: 10.3390/cells9071614
Moran, M., Lynch, C. A., Walsh, C., Coen, R., Coakley, D., and Lawlor, B. A. (2005). Sleep disturbance in mild to moderate Alzheimer’s disease. Sleep Med. 6, 347–352. doi: 10.1016/j.sleep.2004.12.005
Mullington, J. M., Simpson, N. S., Meier-Ewert, H. K., and Haack, M. (2010). Sleep loss and inflammation. Best Pract. Res. Clin. Endocrinol. Metab. 24, 775–784. doi: 10.1016/j.beem.2010.08.014
Murillo-Rodríguez, E., Arias-Carrión, O., Sanguino-Rodríguez, K., González-Arias, M., and Haro, R. (2009). Mechanisms of sleep-wake cycle modulation. CNS Neurol. Disord. Drug Targets 8, 245–253. doi: 10.2174/187152709788921654
Murlidharan, G., Crowther, A., Reardon, R. A., Song, J., and Asokan, A. (2016). Glymphatic fluid transport controls paravascular clearance of AAV vectors from the brain. JCI Insight 1:e88034. doi: 10.1172/jci.insight.88034
Murtha, L. A., Yang, Q., Parsons, M. W., Levi, C. R., Beard, D. J., Spratt, N. J., et al. (2014). Cerebrospinal fluid is drained primarily via the spinal canal and olfactory route in young and aged spontaneously hypertensive rats. Fluids Barriers CNS 11: 12. doi: 10.1186/2045-8118-11-12
Nagelhus, E. A., and Ottersen, O. P. (2013). Physiological roles of aquaporin-4 in brain. Physiol. Rev. 93, 1543–1562. doi: 10.1152/physrev.00011.2013
Neumann, M., Sampathu, D. M., Kwong, L. K., Truax, A. C., Micsenyi, M. C., Chou, T. T., et al. (2006). Ubiquitinated TDP-43 in frontotemporal lobar degeneration and amyotrophic lateral sclerosis. Science 314, 130–133. doi: 10.1126/science.1134108
Ng, A. S., Rademakers, R., and Miller, B. L. (2015). Frontotemporal dementia: a bridge between dementia and neuromuscular disease. Ann. N. Y. Acad. Sci. 1338, 71–93. doi: 10.1111/nyas.12638
Nicaise, C., Soyfoo, M. S., Authelet, M., De Decker, R., Bataveljic, D., Delporte, C., et al. (2009). Aquaporin-4 overexpression in rat ALS model. Anat. Rec. (Hoboken) 292, 207–213. doi: 10.1002/ar.20838
Nielsen, S., Nagelhus, E. A., Amiry-Moghaddam, M., Bourque, C., Agre, P., and Ottersen, O. P. (1997). Specialized membrane domains for water transport in glial cells: high-resolution immunogold cytochemistry of aquaporin-4 in rat brain. J. Neurosci. 17, 171–180. doi: 10.1523/jneurosci.17-01-00171.1997
Nilsson, C., Lindvall-Axelsson, M., and Owman, C. (1992). Neuroendocrine regulatory mechanisms in the choroid plexus-cerebrospinal fluid system. Brain Res. Brain Res. Rev. 17, 109–138. doi: 10.1016/0165-0173(92)90011-a
O’Donnell, J., Zeppenfeld, D., McConnell, E., Pena, S., and Nedergaard, M. (2012). Norepinephrine: a neuromodulator that boosts the function of multiple cell types to optimize CNS performance. Neurochem. Res. 37, 2496–2512. doi: 10.1007/s11064-012-0818-x
Ohene, Y., Harrison, I. F., Nahavandi, P., Ismail, O., Bird, E. V., Ottersen, O. P., et al. (2019). Non-invasive MRI of brain clearance pathways using multiple echo time arterial spin labelling: an aquaporin-4 study. Neuroimage 188, 515–523. doi: 10.1016/j.neuroimage.2018.12.026
Okamoto, Y., Yamamoto, T., Kalaria, R. N., Senzaki, H., Maki, T., Hase, Y., et al. (2012). Cerebral hypoperfusion accelerates cerebral amyloid angiopathy and promotes cortical microinfarcts. Acta Neuropathol. 123, 381–394. doi: 10.1007/s00401-011-0925-9
Ooms, S., and Ju, Y. E. (2016). Treatment of sleep disorders in dementia. Curr. Treat Options Neurol. 18:40. doi: 10.1007/s11940-016-0424-3
Oresković, D., and Klarica, M. (2010). The formation of cerebrospinal fluid: nearly a hundred years of interpretations and misinterpretations. Brain Res. Rev. 64, 241–262. doi: 10.1016/j.brainresrev.2010.04.006
Ouellet, M. C., Beaulieu-Bonneau, S., and Morin, C. M. (2006). Insomnia in patients with traumatic brain injury: frequency, characteristics, and risk factors. J. Head Trauma Rehabil. 21, 199–212. doi: 10.1097/00001199-200605000-00001
Pantazopoulos, H., Woo, T. U., Lim, M. P., Lange, N., and Berretta, S. (2010). Extracellular matrix-glial abnormalities in the amygdala and entorhinal cortex of subjects diagnosed with schizophrenia. Arch. Gen. Psychiatry 67, 155–166. doi: 10.1001/archgenpsychiatry.2009.196
Park, H., Lee, H. K., and Lee, K. (2018). Chronotype and suicide: the mediating effect of depressive symptoms. Psychiatry Res. 269, 316–320. doi: 10.1016/j.psychres.2018.08.046
Patel, T. K., Habimana-Griffin, L., Gao, X., Xu, B., Achilefu, S., Alitalo, K., et al. (2019). Dural lymphatics regulate clearance of extracellular tau from the CNS. Mol. Neurodegener. 14:11. doi: 10.1186/s13024-019-0312-x
Peng, W., Achariyar, T. M., Li, B., Liao, Y., Mestre, H., Hitomi, E., et al. (2016). Suppression of glymphatic fluid transport in a mouse model of Alzheimer’s disease. Neurobiol. Dis. 93, 215–225. doi: 10.1016/j.nbd.2016.05.015
Plog, B. A., Dashnaw, M. L., Hitomi, E., Peng, W., Liao, Y., Lou, N., et al. (2015). Biomarkers of traumatic injury are transported from brain to blood via the glymphatic system. J. Neurosci. 35, 518–526. doi: 10.1523/jneurosci.3742-14.2015
Prasad, A., Bharathi, V., Sivalingam, V., Girdhar, A., and Patel, B. K. (2019). Molecular mechanisms of TDP-43 misfolding and pathology in amyotrophic lateral sclerosis. Front. Mol. Neurosci. 12:25. doi: 10.3389/fnmol.2019.00025
Provenzano, D. A., Pellis, Z., and DeRiggi, L. (2019). Fatal gadolinium-induced encephalopathy following accidental intrathecal administration: a case report and a comprehensive evidence-based review. Reg. Anesth. Pain Med. 44:908. doi: 10.1136/rapm-2019-100422
Rainey-Smith, S. R., Mazzucchelli, G. N., Villemagne, V. L., Brown, B. M., Porter, T., Weinborn, M., et al. (2018). Genetic variation in Aquaporin-4 moderates the relationship between sleep and brain Aβ-amyloid burden. Transl. Psychiatry 8:47. doi: 10.1038/s41398-018-0094-x
Rajna, Z., Mattila, H., Huotari, N., Tuovinen, T., Krüger, J., Holst, S. C., et al. (2021). Cardiovascular brain impulses in Alzheimer’s disease. Brain 144, 2214–2226. doi: 10.1093/brain/awab144
Rangroo Thrane, V., Thrane, A. S., Plog, B. A., Thiyagarajan, M., Iliff, J. J., Deane, R., et al. (2013). Paravascular microcirculation facilitates rapid lipid transport and astrocyte signaling in the brain. Sci. Rep. 3:2582. doi: 10.1038/srep02582
Rasmussen, M. K., Mestre, H., and Nedergaard, M. (2018). The glymphatic pathway in neurological disorders. Lancet Neurol. 17, 1016–1024. doi: 10.1016/s1474-4422(18)30318-1
Reeves, B. C., Karimy, J. K., Kundishora, A. J., Mestre, H., Cerci, H. M., Matouk, C., et al. (2020). Glymphatic system impairment in Alzheimer’s disease and idiopathic normal pressure hydrocephalus. Trends Mol. Med. 26, 285–295. doi: 10.1016/j.molmed.2019.11.008
Reeves, C., Galang, E., Padalia, R., Tran, N., and Padalia, D. (2017). Intrathecal injection of gadobutrol: a tale of caution. J. Pain Palliat. Care Pharmacother. 31, 139–143. doi: 10.1080/15360288.2017.1313353
Ren, Z., Iliff, J. J., Yang, L., Yang, J., Chen, X., Chen, M. J., et al. (2013). ’Hit & Run’ model of closed-skull traumatic brain injury (TBI) reveals complex patterns of post-traumatic AQP4 dysregulation. J. Cereb. Blood Flow Metab. 33, 834–845. doi: 10.1038/jcbfm.2013.30
Ringstad, G., Valnes, L. M., Dale, A. M., Pripp, A. H., Vatnehol, S. S., Emblem, K. E., et al. (2018). Brain-wide glymphatic enhancement and clearance in humans assessed with MRI. JCI Insight 3:e121537. doi: 10.1172/jci.insight.121537
Ringstad, G., Vatnehol, S. A. S., and Eide, P. K. (2017). Glymphatic MRI in idiopathic normal pressure hydrocephalus. Brain 140, 2691–2705. doi: 10.1093/brain/awx191
Roh, J. H., Huang, Y., Bero, A. W., Kasten, T., Stewart, F. R., Bateman, R. J., et al. (2012). Disruption of the sleep-wake cycle and diurnal fluctuation of β-amyloid in mice with Alzheimer’s disease pathology. Sci. Transl. Med. 4:150ra122. doi: 10.1126/scitranslmed.3004291
Román, G. C., Jackson, R. E., Fung, S. H., Zhang, Y. J., and Verma, A. K. (2019). Sleep-disordered breathing and idiopathic normal-pressure hydrocephalus: recent pathophysiological advances. Curr. Neurol. Neurosci. Rep. 19:39. doi: 10.1007/s11910-019-0952-9
Rusting, C. L., and Larsen, R. J. (1998). Diurnal patterns of unpleasant mood: associations with neuroticism, depression, and anxiety. J. Pers. 66, 85–103. doi: 10.1111/1467-6494.00004
Ruz, C., Alcantud, J. L., Vives Montero, F., Duran, R., and Bandres-Ciga, S. (2020). Proteotoxicity and neurodegenerative diseases. Int. J. Mol. Sci. 21:5646. doi: 10.3390/ijms21165646
Samardzic, D., and Thamburaj, K. (2015). Magnetic resonance characteristics and susceptibility weighted imaging of the brain in gadolinium encephalopathy. J. Neuroimaging 25, 136–139. doi: 10.1111/jon.12067
Schain, A. J., Melo-Carrillo, A., Strassman, A. M., and Burstein, R. (2017). Cortical spreading depression closes paravascular space and impairs glymphatic flow: implications for migraine headache. J. Neurosci. 37, 2904–2915. doi: 10.1523/jneurosci.3390-16.2017
Scheuner, D., Eckman, C., Jensen, M., Song, X., Citron, M., Suzuki, N., et al. (1996). Secreted amyloid beta-protein similar to that in the senile plaques of Alzheimer’s disease is increased in vivo by the presenilin 1 and 2 and APP mutations linked to familial Alzheimer’s disease. Nat. Med. 2, 864–870. doi: 10.1038/nm0896-864
Schubert, J. J., Veronese, M., Marchitelli, L., Bodini, B., Tonietto, M., Stankoff, B., et al. (2019). Dynamic (11)C-PiB PET shows cerebrospinal fluid flow alterations in Alzheimer disease and multiple sclerosis. J. Nucl. Med. 60, 1452–1460. doi: 10.2967/jnumed.118.223834
Serrano-Pozo, A., Frosch, M. P., Masliah, E., and Hyman, B. T. (2011). Neuropathological alterations in Alzheimer disease. Cold Spring Harb. Perspect. Med. 1:a006189. doi: 10.1101/cshperspect.a006189
Shea, T. B., and Beermann, M. L. (1994). Respective roles of neurofilaments, microtubules, MAP1B, and tau in neurite outgrowth and stabilization. Mol. Biol. Cell 5, 863–875. doi: 10.1091/mbc.5.8.863
Shokri-Kojori, E., Wang, G. J., Wiers, C. E., Demiral, S. B., Guo, M., Kim, S. W., et al. (2018). β-Amyloid accumulation in the human brain after one night of sleep deprivation. Proc. Natl. Acad. Sci. U.S.A. 115, 4483–4488. doi: 10.1073/pnas.1721694115
Simon, M. J., Wang, M. X., Murchison, C. F., Roese, N. E., Boespflug, E. L., Woltjer, R. L., et al. (2018). Transcriptional network analysis of human astrocytic endfoot genes reveals region-specific associations with dementia status and tau pathology. Sci. Rep. 8:12389. doi: 10.1038/s41598-018-30779-x
Singh, N. N., and Sahota, P. (2013). Sleep-related headache and its management. Curr. Treat. Options Neurol. 15, 704–722. doi: 10.1007/s11940-013-0258-1
Siow, T. Y., Toh, C. H., Hsu, J. L., Liu, G. H., Lee, S. H., Chen, N. H., et al. (2021). Association of sleep, neuropsychological performance, and gray matter volume with glymphatic function in community-dwelling older adults. Neurology 98, e829–e838. doi: 10.1212/wnl.0000000000013215
Sjöholm, L. K., Backlund, L., Cheteh, E. H., Ek, I. R., Frisén, L., Schalling, M., et al. (2010). CRY2 is associated with rapid cycling in bipolar disorder patients. PLoS One 5:e12632. doi: 10.1371/journal.pone.0012632
Stavitsky, K., Neargarder, S., Bogdanova, Y., McNamara, P., and Cronin-Golomb, A. (2012). The impact of sleep quality on cognitive functioning in Parkinson’s disease. J. Int. Neuropsychol. Soc. 18, 108–117. doi: 10.1017/s1355617711001482
Sundaram, S., Hughes, R. L., Peterson, E., Müller-Oehring, E. M., Brontë-Stewart, H. M., Poston, K. L., et al. (2019). Establishing a framework for neuropathological correlates and glymphatic system functioning in Parkinson’s disease. Neurosci. Biobehav. Rev. 103, 305–315. doi: 10.1016/j.neubiorev.2019.05.016
Tabaton, M., and Piccini, A. (2005). Role of water-soluble amyloid-beta in the pathogenesis of Alzheimer’s disease. Int. J. Exp. Pathol. 86, 139–145. doi: 10.1111/j.0959-9673.2005.00428.x
Taoka, T., Jost, G., Frenzel, T., Naganawa, S., and Pietsch, H. (2018). Impact of the glymphatic system on the kinetic and distribution of gadodiamide in the rat brain: observations by dynamic MRI and effect of circadian rhythm on tissue gadolinium concentrations. Invest. Radiol. 53, 529–534. doi: 10.1097/rli.0000000000000473
Taoka, T., Masutani, Y., Kawai, H., Nakane, T., Matsuoka, K., Yasuno, F., et al. (2017). Evaluation of glymphatic system activity with the diffusion MR technique: diffusion tensor image analysis along the perivascular space (DTI-ALPS) in Alzheimer’s disease cases. Jpn. J. Radiol. 35, 172–178. doi: 10.1007/s11604-017-0617-z
Taoka, T., and Naganawa, S. (2020a). Glymphatic imaging using MRI. J. Magn. Reson. Imaging 51, 11–24. doi: 10.1002/jmri.26892
Taoka, T., and Naganawa, S. (2020b). Neurofluid dynamics and the glymphatic system: a neuroimaging perspective. Korean J. Radiol. 21, 1199–1209. doi: 10.3348/kjr.2020.0042
Taoka, T., and Naganawa, S. (2021). Imaging for central nervous system (CNS) interstitial fluidopathy: disorders with impaired interstitial fluid dynamics. Jpn. J. Radiol. 39, 1–14. doi: 10.1007/s11604-020-01017-0
Tarasoff-Conway, J. M., Carare, R. O., Osorio, R. S., Glodzik, L., Butler, T., Fieremans, E., et al. (2015). Clearance systems in the brain-implications for Alzheimer disease. Nat. Rev. Neurol. 11, 457–470. doi: 10.1038/nrneurol.2015.119
Tay, T. L., Savage, J. C., Hui, C. W., Bisht, K., and Tremblay, M. (2017). Microglia across the lifespan: from origin to function in brain development, plasticity and cognition. J. Physiol. 595, 1929–1945. doi: 10.1113/jp272134
Taylor, J. P., Hardy, J., and Fischbeck, K. H. (2002). Toxic proteins in neurodegenerative disease. Science 296, 1991–1995. doi: 10.1126/science.1067122
Toro, C. T., Hallak, J. E., Dunham, J. S., and Deakin, J. F. (2006). Glial fibrillary acidic protein and glutamine synthetase in subregions of prefrontal cortex in schizophrenia and mood disorder. Neurosci. Lett. 404, 276–281. doi: 10.1016/j.neulet.2006.05.067
Torres-Platas, S. G., Nagy, C., Wakid, M., Turecki, G., and Mechawar, N. (2016). Glial fibrillary acidic protein is differentially expressed across cortical and subcortical regions in healthy brains and downregulated in the thalamus and caudate nucleus of depressed suicides. Mol. Psychiatry 21, 509–515. doi: 10.1038/mp.2015.65
Trillo-Contreras, J. L., Toledo-Aral, J. J., Echevarría, M., and Villadiego, J. (2019). AQP1 and AQP4 contribution to cerebrospinal fluid homeostasis. Cells 8:197. doi: 10.3390/cells8020197
van de Haar, H. J., Burgmans, S., Jansen, J. F., van Osch, M. J., van Buchem, M. A., Muller, M., et al. (2016). Blood-brain barrier leakage in patients with early Alzheimer disease. Radiology 281, 527–535. doi: 10.1148/radiol.2016152244
Verkhratsky, A., Nedergaard, M., and Hertz, L. (2015). Why are astrocytes important? Neurochem. Res. 40, 389–401. doi: 10.1007/s11064-014-1403-2
Verma, A., Anand, V., and Verma, N. P. (2007). Sleep disorders in chronic traumatic brain injury. J. Clin. Sleep. Med. 3, 357–362. doi: 10.5664/jcsm.26856
Wang, J., Huang, Q., Li, N., Tan, G., Chen, L., and Zhou, J. (2013). Triggers of migraine and tension-type headache in China: a clinic-based survey. Eur. J. Neurol. 20, 689–696. doi: 10.1111/ene.12039
Watts, R., Steinklein, J. M., Waldman, L., Zhou, X., and Filippi, C. G. (2019). Measuring glymphatic flow in man using quantitative contrast-enhanced MRI. AJNR Am. J. Neuroradiol. 40, 648–651. doi: 10.3174/ajnr.A5931
Webster, M. J., O’Grady, J., Kleinman, J. E., and Weickert, C. S. (2005). Glial fibrillary acidic protein mRNA levels in the cingulate cortex of individuals with depression, bipolar disorder and schizophrenia. Neuroscience 133, 453–461. doi: 10.1016/j.neuroscience.2005.02.037
Williams, M. R., Hampton, T., Pearce, R. K., Hirsch, S. R., Ansorge, O., Thom, M., et al. (2013). Astrocyte decrease in the subgenual cingulate and callosal genu in schizophrenia. Eur. Arch. Psychiatry Clin. Neurosci. 263, 41–52. doi: 10.1007/s00406-012-0328-5
Xia, M., Li, X., Yang, L., Ren, J., Sun, G., Qi, S., et al. (2017a). The ameliorative effect of fluoxetine on neuroinflammation induced by sleep deprivation. J. Neurochem. 146, 63–75. doi: 10.1111/jnc.14272
Xia, M., Yang, L., Sun, G., Qi, S., and Li, B. (2017b). Mechanism of depression as a risk factor in the development of Alzheimer’s disease: the function of AQP4 and the glymphatic system. Psychopharmacology (Berl.) 234, 365–379. doi: 10.1007/s00213-016-4473-9
Xie, L., Kang, H., Xu, Q., Chen, M. J., Liao, Y., Thiyagarajan, M., et al. (2013). Sleep drives metabolite clearance from the adult brain. Science 342, 373–377. doi: 10.1126/science.1241224
Xu, Z., Xiao, N., Chen, Y., Huang, H., Marshall, C., Gao, J., et al. (2015). Deletion of aquaporin-4 in APP/PS1 mice exacerbates brain Aβ accumulation and memory deficits. Mol. Neurodegener. 10:58. doi: 10.1186/s13024-015-0056-1
Yan, T., Qiu, Y., Yu, X., and Yang, L. (2021). Glymphatic dysfunction: a bridge between sleep disturbance and mood disorders. Front Psychiatry 12:658340. doi: 10.3389/fpsyt.2021.658340
Yang, G., Deng, N., Liu, Y., Gu, Y., and Yao, X. (2020). Evaluation of glymphatic system using diffusion MR technique in T2DM cases. Front. Hum. Neurosci. 14:300. doi: 10.3389/fnhum.2020.00300
Yang, W., Wu, Q., Yuan, C., Gao, J., Xiao, M., Gu, M., et al. (2012). Aquaporin-4 mediates astrocyte response to β-amyloid. Mol. Cell. Neurosci. 49, 406–414. doi: 10.1016/j.mcn.2012.02.002
Yang, Y., and Rosenberg, G. A. (2011). Blood-brain barrier breakdown in acute and chronic cerebrovascular disease. Stroke 42, 3323–3328. doi: 10.1161/strokeaha.110.608257
Yin, K. J., Cirrito, J. R., Yan, P., Hu, X., Xiao, Q., Pan, X., et al. (2006). Matrix metalloproteinases expressed by astrocytes mediate extracellular amyloid-beta peptide catabolism. J. Neurosci. 26, 10939–10948. doi: 10.1523/jneurosci.2085-06.2006
Yokota, H., Vijayasarathi, A., Cekic, M., Hirata, Y., Linetsky, M., Ho, M., et al. (2019). Diagnostic performance of glymphatic system evaluation using diffusion tensor imaging in idiopathic normal pressure hydrocephalus and mimickers. Curr. Gerontol. Geriatr. Res. 2019:5675014. doi: 10.1155/2019/5675014
Yuan, H., Lauritsen, C. G., Kaiser, E. A., and Silberstein, S. D. (2017). CGRP monoclonal antibodies for migraine: rationale and progress. Biodrugs 31, 487–501. doi: 10.1007/s40259-017-0250-5
Zbesko, J. C., Nguyen, T. V., Yang, T., Frye, J. B., Hussain, O., Hayes, M., et al. (2018). Glial scars are permeable to the neurotoxic environment of chronic stroke infarcts. Neurobiol. Dis. 112, 63–78. doi: 10.1016/j.nbd.2018.01.007
Zeppenfeld, D. M., Simon, M., Haswell, J. D., D’Abreo, D., Murchison, C., Quinn, J. F., et al. (2017). Association of perivascular localization of aquaporin-4 with cognition and Alzheimer disease in aging brains. JAMA Neurol. 74, 91–99. doi: 10.1001/jamaneurol.2016.4370
Zhang, J., Yang, B., Sun, H., Zhou, Y., Liu, M., Ding, J., et al. (2016). Aquaporin-4 deficiency diminishes the differential degeneration of midbrain dopaminergic neurons in experimental Parkinson’s disease. Neurosci. Lett. 614, 7–15. doi: 10.1016/j.neulet.2015.12.057
Zhang, Z., and Khatami, R. (2014). Predominant endothelial vasomotor activity during human sleep: a near-infrared spectroscopy study. Eur. J. Neurosci. 40, 3396–3404. doi: 10.1111/ejn.12702
Zheng, L., Cedazo-Minguez, A., Hallbeck, M., Jerhammar, F., Marcusson, J., and Terman, A. (2012). Intracellular distribution of amyloid beta peptide and its relationship to the lysosomal system. Transl. Neurodegener. 1:19. doi: 10.1186/2047-9158-1-19
Keywords: glymphatic system, mood disorder, astrocyte, sleep, central nervous system disease
Citation: Zhang D, Li X and Li B (2022) Glymphatic System Dysfunction in Central Nervous System Diseases and Mood Disorders. Front. Aging Neurosci. 14:873697. doi: 10.3389/fnagi.2022.873697
Received: 11 February 2022; Accepted: 31 March 2022;
Published: 25 April 2022.
Edited by:
Ning Song, Qingdao University, ChinaCopyright © 2022 Zhang, Li and Li. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Baoman Li, bmli@cmu.edu.cn